Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Краткие характеристики разных вертикальных зон - Вертикальная зональность - Биогеография морского бентоса - Биогеография бентали Мирового океана
В биогеографии суши, глобальное значение имеет только градиент экватор-полюс, создающий широтную зональность. Вертикальная зональность имеет лишь региональное значение. Это связано с тем, что даже крупнейшие горные системы имеют сравнительно небольшие размеры. В морской биогеографии глобальное значение имеют как и градиент экватор-полюс, и изменение глубины. Причем по последнему градиенту изменения гораздо более существенны, чем по аналогичному градиенту на суше.
“Становится всё более очевидным, что основные подразделения арены жизни в океане — его поверхность и дно, мелководья и абиссальные глубины —имеют принципиальные различия биологической структуры, не укладывающиеся в единую схему биогеографического районирования” (Петров, 2007: 157).
Традиционно схемы районирования бентали по вертикали (глубине) называют схемы вертикальной зональности, и их рассматривают отдельно от схем районирования по “горизонтали”, а схемы районирования по “горизонтали” строят в пределах отдельных вертикальных зон.
Такой подход имел бы право на существование, если бы мощность границ между разными вертикальными зонами была бы существенно больше мощности границ между биогеографическими районами, соседствующими “по горизонтали”. Но это не так. К тому же (1) не существует общепринятой схемы вертикальной зональности (см. ниже); (2) большинство видов обитает в нескольких вертикальных зонах. Напрашивающуюся проблему соотношения выделов разных вертикальных зон практически полностью игнорируют.
На мой взгляд, существование отдельных биогеографий для разных вертикальных зон не имеет под собой никакого обоснования. Но, поскольку почти все предложенные к настоящему времени схемы биогеографии океана сделаны отдельно для разных вертикальных зон, приходится следовать традиции и доступной информации и разобрать вертикальную зональность и горизонтальное биогеографическое районирование отдельно.
Вертикальная зональность
Авторы всех руководств, так или иначе затрагивающих проблемы биогеографического районирования океана, дают схемы вертикальной зональности. Рассмотрим некоторые из предложенных схем.
На Всесоюзном гидрологическом съезде 1928 г. (Рис. 5.43) были выделены следующие вертикальные зоны: супралитораль, литораль, сублитораль (материковое плато), батиаль (материковый склон), абиссаль (ложе океана). Эту схему отечественные авторы в той или иной степени модифицировали (Зёрнов, 1949; Беляев и др., 1959; Беляев, 1966; Виноградова, 1977 и др.) для районирования биоты, хотя схема эта была принята на гидрологическом, а не гидробиологическом съезде. В её основу положены геоморфология и гидрология, а отнюдь не распределение биоты, ещё плохо известное в то время, особенно за пределами шельфа.
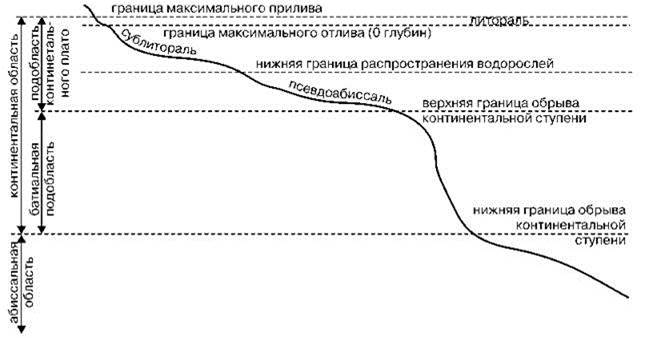
Рис. 5.43. “Схема морских зон” Комиссии Второго Всесоюзного гидрологического съезда. По Аноним (1928) из А.П. Андрияшева (1979).
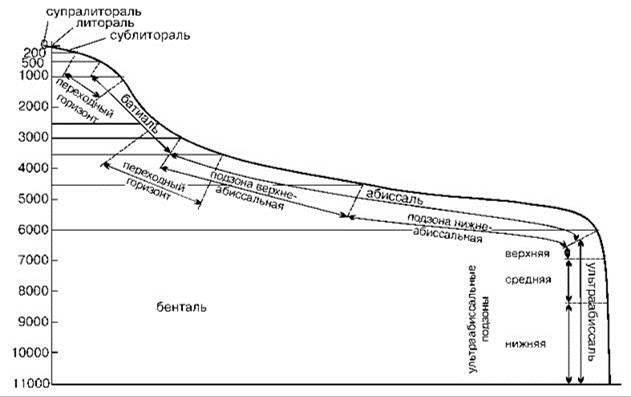
Рис. 5.44. Вертикальная зональность Мирового океана. По Беляеву и др. (1959), Беляеву (1966) из Виноградовой (1977).
Л.А. Зенкевич (1951) дал обзор различных схем вертикальной зональности, начиная с XIX в., и привёл схему, мало отличающуюся от схемы 1928 г. Он выделил литораль (зона приливов-отливов), сублитораль (зона водорослей), псевдоабиссаль (“нижние горизонты эпиконтинентальных водоёмов, если край континентального склона опускается ниже 200 м... в таком случае нижние горизонты континентальной ступени лежат далеко за пределами зоны водорослей, т. е. сублиторали” стр. 294), батиаль (материковый склон) и абиссаль (океаническое ложе).
G.A. Forbes (1854) выделил литораль (<0), зону ламинарий (0—30 м), кораллины (30—90 м) и абиссаль (>100 fms*), глубин (30—90) (цит. по: Danois (1948), 30 и 90 м в оригинале, вероятно, были 15 и 50 fms.
Е. Forbes (1856) выделил литораль (<0), циркумлитораль (0—15 fms), медиаль (15—100 fms) и абиссаль (>100 fms).
S. Ekman (1935, 1953) выделил литораль, архибенталь (от внешней кромки шельфа до примерно 1000 м) и абиссаль — глубины более 1000 м.
A.F. Bruun (1956) выделил шельф, батиаль (внешняя кромка шельфа — 2000 м), абиссаль (2000—6000 м) и хадаль (>6000 м).
J.W. Hedgpeth (1957) выделил шельф, батиаль (внешняя кромка шельфа — 4000 м), абиссаль (4000—6000 м) и хадаль (>6000 м).
В Институте океанологии была разработана схема вертикальной зональности (Беляев и др., 1959; Беляев, 1966), включающая супралитораль, литораль, сублитораль (0—200 м или до края шельфа), батиаль (материковый склон от 200 м или края шельфа до 3000 м), абиссаль (океаническое ложе, 3000—6000 м) и ультраабиссаль (глубоководные желоба с глубинами более 6000 м) (Рис. 5.44).
А.С. Константинов (1972) выделил супралитораль, литораль, сублитораль (0 — “нижняя граница распространения донных фотосинтезирующих растений” или 200 м), батиаль (материковый склон), абиссаль (океаническое ложе), ультраабиссаль (>6—7 км).
J.C. Briggs (1995) выделил шельф (<200 м), склон (200—2000 м), абиссаль (2000—6000 м) и хадаль (>6000 м).
А.И. Кафанов, В.А. Кудряшов (2000: 75) различали “зону заплеска и штормовых выбросов — супралитораль, приливно-отливную зону, или литораль, зону материковой отмели, или шельфа — сублитораль, зону континентального склона — батиаль, зону океанического ложа — абиссаль, зону глубоководных океанических желобов — ультраабиссаль, или хадаль”.
“Резкой границы между литоралью [sic! — ИЖ] и абиссалью нет, их разделяет переходная батиальная зона” (Абдурахманов и др., 2001: 307).
Легко заметить, что набор вертикальных зон остаётся примерно постоянным, особенно в последние полвека: супралитораль, литораль, сублитораль (подразделяемая иногда на несколько зон), батиаль, абиссаль и ультраабиссаль. Варьируют лишь глубины, по которым проводят границы зон и их названия. Так, в нашей стране наиболее глубокую зону называют ультраабиссаль, а за рубежом — хадаль. Причина этого состоит в том, что изначально полагали: жизни на таких глубинах нет. В 1948 г. во 2-м рейсе “Витязя” с глубины 8100 м в на станции 162 (44°55,5 N 152°24 E) подняли трал, полный полихет, которых П.В. Ушаков определил, как Amphicteis mederi. Открытие жизни на таких глубинах тут же засекретили. Позже в рейсе датского научно-исследовательского судна “Galathea” также была открыта жизнь на глубинах более 6000 м, и датчане написали об этом Л.А. Зенкевичу. Он тут же пошёл в КГБ и просил разрешить публикацию для закрепления приоретета советских учёных. Публикацию разрешили. Но было уже поздно: публикация датчан вышла на несколько месяцев раньше. Российские и датские учёные предложили разные термины для обозначения вертикальной зоны глубоководных желобов, что и закрепилось в мировой практике.
Перечисленные выше зоны в биогеографическом отношении неравноценны. Литораль, по крайней мере литораль умеренных широт, и, тем более, супралитораль, нельзя рассматривать как биогеографический выдел. Практически все литоральные виды морского происхождения обитают преимущественно или исключительно на литорали из-за биотического взаимодействия. В ЦС с пониженным таксономическим разнообразием литоральные виды способны обитать и в сублиторали, часто чувствуя себя там гораздо лучше. Несколько примеров:
— В Балтийском море, биота которого очень обеднена из-за пониженной солёности, типично литоральные бурые водоросли Fucus vesiculosus и Ascophyllum nodosumобитают до глубины 20 м (Sergerstrele, 1957).
— Начиная с плейстоцена, у бореальных атлантических берегов Северной Америки практически отсутствуют участки с каменистым дном на литорали и верхней сублиторали. Поэтому соответствующие сообщества там сильно обеднены и фактически образованы лишь частью видов, обитающих в сходных биотопах у берегов Европы, свои же виды из-за редкости биотопа не образовались. Поэтому такие брюхоногие моллюски, как Littorina littorea, L. “saxatilis” и Thais lappilus, у берегов Европы обитающие по большей части на скалистой литорали, у берегов Северной Америки обычны на каменистых грунтах и верхней сублиторали (Vermeij, 1978).
Иными словами, литораль — это биотоп, населённый малоконкурентоспособными верхнесублиторальными видами, обитающими в неоптимальных для них условиях. Граница между литоралью и сублиторалью — это граница между сообществами одной ЦС, а не биогеографическая, поскольку нет эндемов литорали, между литоралью и сублиторалью нет границ ареалов. Некоторым аналогом литоральных видов может служить экологическая группа растений, произрастающих в сильно затенённых местах. Детальный анализ их биологии давно привёл ботаников к выводу, что нет тенелюбивых растений, а есть теневыносливые, причём для большей части видов лимитирующим абиотическим фактором, ограничивающим их распространение в освещённых участках, является не избыток света, а недостаточная увлажнённость. Виды, обитающие на литорали, можно аналогично назвать “литоралевыносливыми”, и для большинства из них специфический режим литорали (чередование приливов и осушения) вовсе не является необходимым, но они способны его выносить. Схемы биогеографического районирования литорали поэтому имеют совсем другой смысл, нежели биогеографическое районирование других вертикальных зон (сублиторали, батиали, абиссали и др.).
Исключение составляет лишь верхняя литораль низких широт, где развиваются весьма специфические ассоциации и в первую очередь мангры. Состав мангровых ассоциаций позволяет рассматривать их как в составе наземных СС, так и делать предметом морской биогеографии. Ниже они для простоты рассмотрены в составе верхнесублиторальных сообществ, хотя эта точка зрения, возможно, ошибочна.
Первые схемы вертикальной зональности были, по всей видимости, просто представлениями экспертов. Позднее положение границ между вертикальными зонами устанавливали с использованием различных методов. В нашей стране наибольшее распространение получил метод биотических разрезов, и соответственно получаемые результаты имеют все свойственные методу недостатки (подробнее см. Главу 2). В частности, данные по распространению отдельных видов по площадям различных размеров (от сравнительно небольших участков до всего Мирового океана) проецируют на ось глубин. При этом неизбежно, как и при любом проецировании трёхмерного объекта на одну ось, происходит искажение действительной биогеографической картины, причём всегда в одну сторону — в сторону размывания границ, приводящее к утрате резкости изменений таксономического состава на границе, что на графике приводит к сглаживанию пиков. Критерий, по которому пик считают достаточным, чтобы быть границей, субъективен. Даже если оценивают различия списков биот по разные стороны границы математически, критерий различия субъективен — 5%, 1% и т. п.
“Как раньше, так и в настоящее время вертикальное членение донной фауны проводилось по основным структурам дна, ... всегда имелись в виду и другие факторы, особенно более резкие градиенты в их распространении” (Шунтов, 2001: 400).
Но основная проблема даже не в методе, а в первоначальной разбивке оси глубин на диапазоны для сравнения, а также в интерпретации результатов. Если в качестве основания для разбивки вертикальной оси используют геоморфологию или глубину, то изучают не распределение биоты в пространстве, а различия геоморфологических структур или глубин, т. е. используют географический, а не биологический подход. Бессмысленно в этом случае говорить о совпадении распределения биоты с геомофологией или её приуроченности к определённым глубинами, если их изначально использовали для разделения отрезка изучаемой оси, поскольку это совпадение предопределено методически.
При использовании данных геоморфологии получаемая схема вертикальной зональности неизбежно дублирует геоморфологическую, а термины сублитораль, батиаль, абиссаль и ультраабиссаль становятся излишними синонимами геоморфологических. За рубежом, кстати, термин “сублитораль” почти и не используют, обходясь понятием “shelf”, часто и вместо “батиали” используют “slope”. Но, поскольку крупномасштабная вертикальная зональность, по-видимому, действительно связана с геоморфологией, районирование, полученное на основе выделения зон по геоморфологическим критериям, вероятно, отражает, хотя и с большими или меньшими искажениями, реальное распределение ЦС по глубине. Искажения возникают и от того, что прекрасно смотрящиеся на схеме морфологические структуры (шельф, склон, ложе и т. п.) в реальности совсем не такие: увеличение глубин вовсе не монотонно, склон прорезают каньоны, часто далеко вдающиеся в шельф и имеющие у своего подножия на ложе конус выноса и т. д. и т. п. Границы шельфа, склона, ложа и ультраабиссальных котловин расположены на различных глубинах в разных, порой соседних районах. При составлении же схем вертикальной зональности методом биотических разрезов всё это вынужденно (из-за малого числа данных) игнорируют. Кроме геоморфологии на распределение видов также влияет распределение водных масс, которое, хотя и связано с геоморфологией, но всё же их связь далека от функциональной.
Если же зоны выделяют по глубинам, то часто получают схему, которая вовсе ничему не соответствует. Например, если взяты интервалы 0—200 и 200—1000 м, то смена таксономического состава по результатам анализа не может происходить нигде, кроме как на глубине 200 м, которую и принимают за глубину прохождения биогеографической границы. Между тем, она может проходить в данном месте на глубине, например, 163 м. То, что подсчёт вели для другого диапазона глубин, приводит не только к неправильному определению положения границы, но и к уменьшению её чёткости, поскольку при построении графика в состав биоты верхнего горизонта были включены и виды нижнего (так как обитание их на глубинах 163—200 м вполне нормально).
Особо надо сказать о глубине 200 м, используемой во многих схемах. 200 м — геоморфологическое понятие, оно означает, что данное место находится на 200 м глубже уровня Мирового океана. Что же есть такого любопытного на этой глубине? Часто говорят, что это — глубина кромки шельфа. Но это не так, на самом деле средняя глубина кромки шельфа в Мировом океане — 137 м (Шипард, 1969). Однако таковой является средняя глубина, но в отдельных районах кромка шельфа располагается на совсем других глубинах. Например, у берегов южного Перу и северного и центрального Чили край шельфа находится на глубине 50 м (Удинцев, 1972; Семёнов, 1982: 255). В приполярных районах (Северный Ледовитый океан, Антарктика) кромка шельфа, наоборот, располагается заметно глубже, что объясняют бывшими здесь оледенениями. Многокилометровая толща ледника своим весом чуть притопила материк, после таяния ледника он медленно всплывает (скорость подъёма в Осло- фьорде — 1,5 см/год (Бьёрке, 1993). У прежнего центра оледенения Скандинавии (в вершине Ботнического залива) подъём за последние 9000 лет составил 250 м, но процесс ещё не завершён, поскольку магматические породы (базальты и др.) — плохо текущая жидкость, и уровень земли поднимется, по-видимому, в данном районе ещё на 200 м (Монин, 1977). Впрочем, — это не единственная, и, вероятно, не основная причина столь глубокого шельфа, поскольку у северных берегов Аляски, где располагался ледник значительно больший, чем в Скандинавии, кромка шельфа расположена на глубине всего 70 м (Carsola, 1954; Carsola et. al, 1961). Кроме того, ледник при движении производит разрушение подстилающей поверхности Земли (ледниковая абразия). Образованный материал ледник переносит к краю и отлагает в виде валов (морен) на границе ледника, а в случае плавучих ледников — на границе всплытия (там, где лежащий на грунте ледник переходит в плавучий). В результате абразивного действия ледника подстилающая его поверхность становится ниже, а прилегающая часть материкового склона — выше. Так, по имеющимся данным (Гроссвальд, 1983; Матишов, 1986) всё дно Баренцева моря подвергалось ледниковой абразии, и краевые морены расположены вдоль кромки шельфа. В результате, хотя с геоморфологической точки зрения Баренцево море расположено целиком на шельфе, его максимальная глубина — 600 м, и даже средняя — 222 м (Добровольский, Залогин, 1982), т. е. по критерию “200 м” оно вообще не является шельфом. Таким образом, глубину кромки шельфа определяют геологические процессы и закон Архимеда, но они ничего не знают о цифре “200 м”.
Л.А. Зенкевич (1951: 196) считал 200 м нижней границей распространения донных водорослей, но и это не так, они обнаружены и на вдвое больших глубинах, но, главное, уже на глубине в несколько десятков метров их роль столь незначительна, что выделять на этой основе зону неоправданно.
Другой возможный смысл числа “200 м” — “глубина, глубже которой самые крупные океанские волны уже не могут взмучивать самые мелкие частицы наносов” (Морская геоморфология, 1980). В реальном океане задолго до того, как перестаёт сказываться влияние волн на осадки, перенос осадков волнами становится незаметным на фоне переноса осадков придонными течениями, вызванными иными причинами. Таким образом, и это значение цифры “200 м” незначимо с точки зрения биологии.
Еще одна цитата: “В наши дни глубина 200 м является предельной для широкого экономически выгодного применения существующих технических средств поиска и добычи биологических и минеральных богатств моря” (Баскаков и др., 1987: 17). И это значение не имеет отношения к биологии.
“Глубины моря являются чрезвычайно разнообразными; онъ выражаются, обыкновенно, въ морскихъ саженяхъ” (Келлер, 1897: 28).
По-видимому, существует только одно сколько-нибудь разумное объяснение широкого использования данного числа: до XIX в., а в некоторых странах и сейчас, расстояния в море (и глубину в том числе) мерили в морских милях и саженях. В схеме Е. Forbes (1856) граница между медиальной и абиссальной зонами проходит по глубине 100 fms. 100 морских саженей — психологически осмысленная величина, тем более, что в то время знания о населении таких глубин были очень незначительны. Эта схема была длительное время широко распространена, и современные схемы вертикальной зональности являются её развитием. С переходом на метрическую систему мер привычные 100 fms было неудобно заменять на 182,5 м, поэтому округлили до 200 м. В Англии, где метрическая система мер до сих пор не стала единственной, до сих пор 200 м от 100 fmsиспользуют как равноправные величины. В частности, в большинстве выпусков аналога “Фауны России” — British Synopses (серийное издание определителей, как всякая серия с унифицированным оформлением, содержанием и, конечно, рассматриваемой в отдельных выпусках акваторией) рассматриваемая акватория ограничена изобатой 200 м (см. например, Southward, Campbell, 2006: fig.1), однако в некоторых (например: Cornelius, 1995: fig.1) изобатой 600 футов (= 100 fms = 183 м).
Ясно, что при таком происхождении величины “200 м”, в ней не больше смысла, чем в семи верстах, которые надо пройти, чтоб похлебать киселя. Поэтому использовать 200 м для выделения каких-либо биологически осмысленных вертикальных зон более чем странно.
Можно отметить, что в странах, где метрическая система не является единственной, используют и другие глубины кромки шельфа. Так Kaiser et al. (2005), Woodward (2008) используют глубину 500 футов (=152,4 м), что гораздо ближе к средней глубине кромки шельфа, но также не является биологической величиной.
Всякий термин надо определять через свойства объектов, к которым он относится, т. е. биологический термин надо определять через свойства биологических объектов. Поэтому, если уж использовать термины сублитораль, батиаль и абиссаль, то надо вкладывать в них иной, не геоморфологический, а гидробиологический смысл и понимать под ними биогеографические регионы, сменяющие друг друга по глубине.
Нельзя не упомянуть схему А.П. Андрияшева (1979) (Рис. 5.45). Она является развитием схемы вертикальной зональности 1928 г. (Рис. 5.43). Положительными чертами его схемы является разделение сублиторали на верхнюю (= сублитораль s.str.) и нижнюю, границу между которыми он проводит по нижней границе пояса макрофитов и рифообразующих кораллов. С этим следует согласиться, уточнив лишь, что не всех рифообразующих кораллов, а герматипных. Дав исторический обзор использования термина “псевдоабиссаль”, А.П. Андрияшев указывает на очевидную его непоследовательность и предлагает полностью изменить его смысл и вводит термин “псевдобатиаль”. Он определяет псевдобатиаль как “сравнительно небольшие локальные котловины в пределах шельфа (как, например, в Белом море или на антарктическом шельфе)”. Псевдоабиссаль — “большие глубоководные, частично изолированные моря вроде Средиземного или Японского” (Андрияшев, 1979: 123). Судя по приведённым определениям, логическим основанием для обоих терминов предстают глубина и рельеф, поэтому они являются геоморфологическими терминами, а не биологическими. Фактический их смысл — повторение части информации обычной батиметрической карты. Термины же батиаль и абиссаль — хорологические, относящиеся к специфическим биотам. Использование геоморфологических терминов, образованных от корней хорологических, без нужды путает и без того запутанный предмет. Поэтому я считаю их использование не только излишним, но и вредным.

Рис. 5.45. Схема вертикальной зональности по А.П. Андрияшеву (1979).
Примечание. Нижняя граница распространения рифообразующих кораллов находится на глубине 1—2 км и отнюдь не совпадает с нижней границей пояса макрофитов; по-видимому, А.П. Андрияшев имел в виду облигатно герматипные кораллы.
Также А.П. Андрияшев вводит термин талассобатиаль для батиальных районов океанических островов и подводных гор, удалённых от материков. Используя этот термин, надо помнить об отсутствии резких различий между обычной батиалью и талассобатиалью. Степень изоляции разных районов талассобатиали от обычной батиали варьирует от очень сильной до практически отсутствующей. В общем, для талассобатиали характерны рассмотренные выше особенности островных биот: одни районы талассобатиали отличаются высокой специфичностью биоты, другие — практически неотличимы от обычной батиали.
Изучение распределения видов по глубине, а не сравнение по видовому составу районов, выделенных по небиологическим критериями (различных глубин или разных крупных геоморфологических структур), даёт картину, существенно отличную от классической, причём разную в разных районах. Имеющиеся данные по вертикальному распределению бентоса показывают, что смена биот происходит на различных глубинах в разных районах Мирового океана (Menzies et al., 1973; Миронов, 1986) (Рис. 5.46), и их распространение примерно соответствует макрорельефу дна (а не глубине). Причина этого в том, что именно с макрорельефом дна связаны характер распределения взвеси в придонных слоях воды и характер осадконакопления. Кроме макрорельефа смену вертикальных зон определяют и водные массы: верхняя сублитораль расположена в прибрежных водных массах, для которых характерно наличие освещения, достаточного для фотосинтеза, сезонные колебания гидрологических показателей (температуры в холодных, умеренных и субтропических водах и солёности в тропиках) и другие особенности (см. выше). Иногда расположение водных масс определяет вертикальную зональность и глубже. Это наблюдается, например, у Скандинавии и в Охотском море. Распределение водных масс одновременно связано и с рельефом, особенно такое относится к водным массам, расположенным выше главного термоклина. Необходимо помнить, что при увеличении размера анализируемой акватории, методически неизбежно будет происходить уменьшение чёткости границ, а некоторые могут вообще исчезнуть.
Сублитораль ранее (Forbes, 1856; Всесоюзный гидрологический съезд, 1928; Зенкевич, 1951) разделяли на верхнюю и нижнюю (Рис. 5.43). Но примерно с середины прошлого века (Ekman, 1935, 1953 и др.) её стали рассматривать как единую зону. Какого-либо серьёзного обоснования такой смены точек зрения в литературе я не встречал. В то же время, при анализе достаточного количества материала неизбежно оказывалось, что шельф неоднороден по вертикали, а положение границ между вертикальными зонами и даже их число варьирует. Приведу несколько примеров, показывающих, что верхняя и нижняя сублитораль населены различными биотами = между верхней и нижней сублиторалью проходят границы ареалов видов.
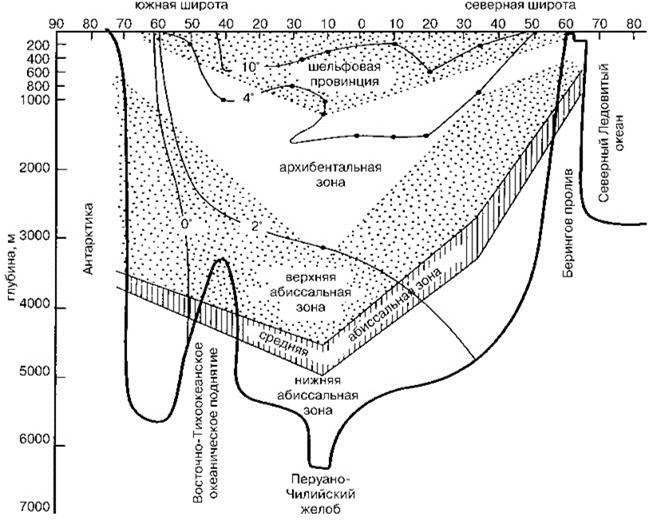
Рис. 5.46. Глубина залегания вертикальных зон вдоль меридионального разреза в Тихом океане. По Menzies et al. (1973).
В.В. Федяков (1986) показал, что фауна поверхностной и нижней водных масс (глубины варьируют в широких пределах в зависимости от места) на разных участках Белого моря очень сильно различаются соотношением видов с различным географическим распространением.
По данным А.И. Кафанова (1991), в Северной Пацифике в целом и в отдельных её районах фауна двустворчатых моллюсков прибрежных мелководий отличается от фауны открытого шельфа на уровне отчётливого биогеографического выдела. Так, он пишет (стр. 70—71): “стенобатные литорально-верхнесублиторальные виды составляют не менее 40% от их общего числа” (видов, встречающихся на глубинах до 200 м — ИЖ), т. е. даже по формальному критерию S.P. Woodward (1856) литораль + верхняя сублитораль и нижняя сублитораль являются самостоятельными биогеографическими регионами, поскольку происходит смена половины биоты. Несмотря на это, сам А.И. Кафанов считает различие биогеографически незначимым.
О.А. Скарлато (1982) отмечает, что видовой состав двустворчатых моллюсков закрытых бухт отличается от видового состава открытых мысов в той же степени, что и выделяемые им провинции. У открытых мысов прибрежный фронт расположен глубже и большая часть исследованной в работе акватории находится в прибрежной водной массе, поэтому различия мысов вероятно объясняются различиями биот верхней и нижней сублиторали.
Г.М. Каменев (1990, 1995) показал, что у Командорских островов по распространению двустворчатых моллюсков можно выделить три вертикальные зоны: литораль — 20 м, 60—100 м и глубже 130—200 м, причём сходство моллюсков на глубинах 0—20 м и >40 по индексу Чекановского-Сёренсена 45%.
П.А. Балыкин (2007) отмечает, что фауны рыб западной части Берингова моря на глубинах <100 м и 100—200 м имеют только 64% общих видов.
О.Г. Кусакин и С.А. Ростомов (1982) показали, что фауны Isopoda на глубинах 0—5 м и более 250 м во всех трёх исследованных ими районах вообще не имеют общих видов (Рис. 5.47). Тем не менее, они отнесли данные районы к одному биогеографическому региону. При этом эндемичные для верхних и нижних горизонтов шельфа группы видов они посчитали низкобореальными несмотря на то, что в двух районах они даже не встречаются вместе. Чтобы различить мелководные и глубоководные низкобореальные виды, авторы использовали отношение видов к температуре — логическое основание, использование которого делает классификацию не биогеографической.
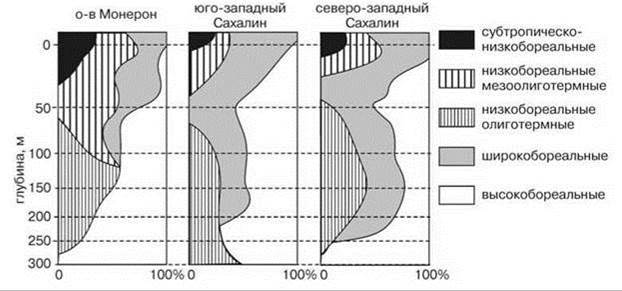
Рис. 5.47. Вертикальное распределение разных географических элементов на шельфе севера Японского моря. По О.Г Кусакину и С.А. Ростомову (1982).
А.П. Андрияшев (1939: 24) выделял в ихтиофауне Берингова моря:
— Виды прибрежных мелководий, обитающие большей частью на глубинах от литорали до 15—20 м, а зимой часто глубже (т.е. виды прибрежной водной массы, граница которой зимой располагается на больших глубинах).
— Верхнесублиторальные виды, обитающие на глубинах от 20—30 до 100—120 м.
— Нижнесублиторальные виды, обитающие на глубинах от 100—120 м до 250—300 м. Среди последних двух групп видов имеются викарирующие виды.
— Глубоководные виды, обитающие глубже 250—300 м.
К.В. Бандурин (2007), анализируя распространение креветок на шельфе севера Охотского моря, выделяет три типа ареалов: литорально-верхнесублиторальные, сублиторальные (преимущественно до 100 м) и шельфовые (преимущественно на 200-300 м).
А.Н. Голиков указывает, что “северная граница Японской низкобореальной подобласти проходит в районе м. Терпения и северной части о. Итуруп. Однако глубины свыше 30 м в заливах Анива и Терпения занимает Алеутская высокобореальная подобласть” (Голиков, 1982: 97). Возможно, пытаясь отразить неоднородность биот по глубине, но, оставаясь в рамках “биогеографии всего шельфа”, А.Н. Голиков (1980, 1982) вводит в биогеографическую характеристику отдельных видов данные по глубине их обитания, что при выделении действительно биогеографических регионов было бы совершенно излишне.
Надо подчеркнуть, что широко распространенная точка зрения, согласно которой на шельфе расположена только одна вертикальная зона, не является, однако, общепризнанной. Так, разделить на биогеографическом уровне население верхней сублиторали и глубже расположенных частей шельфа ранее предлагала на основе анализа распространения десятиногих раков залива Анива (южный Сахалин, Охотское море) З.И. Кобякова (1956). Аналогичные фаунистические различия в пределах шельфа имеются и в бореальных районах европейских вод, а также и в тропическом Индо-Малайском архипелаге (Миронов, 1986). Ю.Е. Петров (1977) выделяет на шельфе верхнюю (до 6—10, реже 25 м) и нижнюю (до 40—50, реже 200 м) сублитораль.
Тем не менее, большинство исследователей считает сублитораль единой вертикальной зоной, соответствующей шельфу. Возможная причина этого состоит в том, что бентосные работы с судов не охватывают мелководье (верхнюю сублитораль), а с берега — большие глубины (нижнюю сублитораль). Обработку собранных с берега и с судов материалов проводят обычно отдельно. И в обоих случаях нет особых причин сомневаться в их биогеографической неоднородности. Объединение же данных береговых и судовых работ проводят с уже сформировавшимся мнением об отсутствии биогеографических различий верхней и нижней сублиторали.
“Литоральная фауна северных берегов Франции и Северного моря идёт на север и восток до Белого моря, мало меняя свой основной состав, но существенно меняется в южной части атлантического побережья Франции. Сублиторальная фауна Северного моря на север доходит только до северных частей Скандинавии, но в нижних слоях мало прогреваемой воды идёт гораздо дальше на юг” (Зенкевич, 1946: 123).
Другая возможная причина — распространённое представление о наличии разных масштабов: одни явления объявляют мелкомасштабными, а другие крупномасштабными, схемы же вертикальной зональности считают схемами крупномасштабных структур. Однако, никаких биологических критериев для пороговых значений при определении “масштабности” в настоящее время не предложено, этот вопрос фактически не изучен. Некоторые подходы к его решению намечены в докторской диссертации А.И. Азовского (2002), но проблема далека от окончательного решения. Единственный используемый в настоящее время критерий — субъективный, что, как минимум, позволяет существовать обеим точкам зрения. В то же время, масштабы верхней сублиторали являются малыми только в направлении, перпендикулярном берегу. Вдоль же берега размеры соответствующих регионов имеют протяженность сотни и тысячи километров, что никак нельзя считать мелкомасштабным явлением. В данном случае мы имеем дело с ленточными структурами-явлением вовсе не уникальным для верхних отделов шельфа. Подобные структуры сходных и даже меньших размеров встречаются и в других местах, при этом биогеографы (иногда те же самые) считали возможным рассматривать их как самостоятельные биогеографические регионы:
— Батиаль. Протяженность обычно признаваемой в качестве самостоятельного биогеографического региона батиали в направлении, перпендикулярном кромке шельфа, также весьма невелика; она сопоставима с верхней сублиторалью (обычно 15—30 км).
— Ультраабиссаль. Ширина утраабиссальных желобов по изобате 6000 м (часто принимаемой за границу ультраабиссали) редко превышает 30 км (до 200 км), а по дну не превышает нескольких десятков км.
— Талассобатиаль. Биоты гор и изолированных поднятий часто рассматривают как самостоятельные биогеографические выделы (Андрияшев, 1979; Миронов, 1994; Парин, Миронов, Несис, 1997; Миронов, Крылова, 2006 и др.), хотя их линейные размеры часто составляют десятки километров или даже километры.
— Вертикальная зональность в горах. А.И. Толмачев при выделении конкретных флор считает необходимым “учитывать особенности поясной дифференциации её растительного покрова, наличие приуроченных к определённым уровням флористических рубежей” (Толмачев, 1974: 188). В частности, при изучении флор Гиссаро-Зеравшанской горной страны “выявляется наличие настолько резко выраженного рубежа между флорами альпийского и субальпийского поясов, что более правильным кажется обособленное рассмотрение альпийской флоры конкретных частей этого высокогорья” (Толмачев, 1974: 188).
Поскольку размеры этих ленточных структур примерно те же, что и у верхней сублиторали, очевидно, что сам по себе размер структуры не является сколь-нибудь значимым для того, считать ли её биогеографической единицей или нет. Только традицией можно объяснить, почему в одном случае ленточную структуру одного размера объявляют мелкомасштабной, а в другом структуру того же размера — нет. Об этом, кстати, свидетельствует то, что, не выделяя верхнюю сублитораль в самостоятельный биогеографический выдел, некоторые авторы, например, А.И. Кафанов и В.А. Кудряшов (2000) выделяют гидротермальную область, хотя размеры отдельных гидротерм в любом направлении куда как меньше размеров верхней сублиторали даже в направлении, перпендикулярном берегу.
Граница между верхней и нижней сублиторалью примерно соответствует положению прибрежного фронта. Примерно, поскольку положение прибрежного фронта определяет гидрология. Динамика гидрологических условий гораздо выше динамики бентоса: большая часть организмов макробентоса обладает малой подвижностью и продолжительностью жизни, превышающей год. Глубина, над которой расположен прибрежный фронт пропорциональна кубу скорости течения на поверхности (см. выше), т. е. она не только меняется в течение года, но и существенно уменьшается с уменьшением скорости течений (гораздо меньше в бухтах, чем у открытых мысов). Поэтому изучение распределения бентоса в зависимости только от глубины неизбежно “размывает” эту границу, создавая обманчивое впечатление плавного перехода верхнесублиторальной биоты в нижнесублиторальную.
Детальный анализ распределения зоопланктона также показал, что следует разделять сообщества прибрежной водной массы и вод открытого шельфа (Шунтов, 2001: 214).
Сколько же всего вертикальных зон и на каких глубинах они примерно расположены?
“Несомненным оказываются лишь два факта: широкая эврибатность большей части фауны и полное отсутствие специфической сублиторальной и вообще мелководной фауны <Антарктики>” (Грузов, 1980: 29—30).
В Северном Ледовитом океане (в высокой Арктике), по-видимому, всего две вертикальные зоны: шельфовая (без подразделения на верхнесублиторальную и элиторальную) и глубоководная (без подразделения на батиальную и абиссальную), ультраабиссальная зона отсутствует из-за недостаточно больших глубин. Отсутствует верхняя сублитораль и у Антарктиды. Вероятные в обоих случаях причины: отсутствие пребрежного фронта и то, что верхние горизонты шельфа были покрыты ледником в плейтоцене. Некоторым аналогом верхней сублиторали в Северном Ледовитом океане можно считать распреснённые участки морей. Обильный сток в Северный Ледовитый океан (превосходящий сток в Индийский) при небольшом размере самого океана (4% Мирового океана) делает участки с пониженной солёностью обычными и занимающими большую площадь. Многие обитающие в Арктике виды обильны в распреснённых районах и почти отсутствуют в акваториях с нормальной солёностью. Однако солёность в распреснённых районах варьирует в значительных пределах (от пресной до морской), как от места к месту, так и во времени. Причём варьирование это не только значительно, но и мало предсказуемо. Это позволило сформироваться в распреснённых районах лишь эвригалинным видам, но не ЦС. Таксоны, которые не смогли приобрести эвригалинность избегали распреснения, уходя на большие глубины. Поэтому эндемичные арктические виды или эвригалинны, или эврибатны.
Н.Н. Детинова (1993), изучая вертикальное распределение полихет у Намибии, в диапазоне 124—1155 м выделила три зоны, границы между которыми расположены на глубине 300 м и 500—700 м. Зона, расположенная на глубине от 300 до 500—700 м, несмотря на узость, имеет даже более высокое абсолютное число эндемов, чем ниже и вышележащие зоны. Верхняя сублитораль её исследованиями не была охвачена, поэтому можно утверждать, что у Намибии четыре вертикальные зоны на шельфе и верхней части склона.
Р.Н. Буруковский (1984), изучая вертикальное распределение креветок у Западной Африки, выделил на глубинах 13—1230 м четыре самостоятельные вертикальные зоны, каждой из которых соответствует свой таксоцен креветок: шельфовый (до 50—100 м), кромки шельфа (от примерно 70 м до 300 м и более), верхней (примерно 300—800 м) и нижней части склона (глубже 700—800 м). Этот результат соответствует зональности у Намибии.
О.Н. Зезина (1973), анализируя вертикальное распространение брахиопод в Мировом океане, выделяет между нижней сублиторалью и батиалью горизонт с эндемичной фауной.
Olabarria (2004), анализируя вертикальное распространение двустворчатых моллюсков в северо-восточной Атлантике на глубинах 500—4866 м, выделяет зоны: выше 750 м, переходную от шельфа к батиали, верхне- и среднебатиальную 750—1900 м, нижнебатиальную 1900—2900 м и абиссальную глубже 4100 м, на глубинах 2900—4100 м расположена переходная зона между батиалью и абиссалью, где обитают и батиальные, и абиссальные виды.
В National Museum of Wales (Cardiff, UK) принята схема: литораль, континентальный шельф (до 200 м), зона края континента (200—500 м), батиаль (500—2000 м), абиссальный склон (2000—4000 м), абиссальная равнина (>4000 м) (Oliver et al., 2010).
А.М. Орлов (2004) на основе анализа распределение рыб в нижней части шельфа и верхней части склона у тихоокеанского побережья Курильских островов и юго-восточной Камчатки выделил сублитораль, элитораль, мезобенталь и батибенталь. Границы между тремя последними зонами ними он провёл, соответственно, на 150—200 м и 500—550 м.
Число подобных примеров можно продолжить, но очевидно, что даже общее число вертикальных зон, а тем более конкретные глубины, на которых проходят границы вертикальных зон, в разных районах варьируют, а, следовательно, процесс составления схемы вертикальной зональности весьма далёк от своего завершения. Тем не менее, не только авторы многих учебников, но и научных работ, часто пытаются свести сложную картину вертикальной зональности к четырём, а то и двум-трём зонам. Здесь уместно ещё раз напомнить едкое высказывание Ф. Дарлингтона.
“Если зоогеограф опирается на авторитет мнений, это означает, что у него не хватает фактов или что он ошибается” (Дарлингтон, 1966: 32).
Типичным набором вертикальных зон, по-видимому, пока можно считать следующий (Рис. 5.48): (1) верхняя сублитораль (сублитораль в узком смысле), включая литораль, (2) элитораль, или нижняя сублитораль, (3) верхняя батиаль = зона кромки шельфа, (4) нижняя батиаль, (5) абиссаль и (6) ультраабиссаль, или хадаль. Набор этот типичный, но отнюдь не везде вертикальных зон именно шесть. В некоторых случаях, возможно, на склоне расположено больше двух зон. В тоже время, в некоторых случаях зона кромки шельфа, возможно, является не самостоятельной зоной, а экотоном, специфичность биоты которого обусловлена пограничным эффектом. С другой стороны, в приполярных районах, как уже было показано, по-видимому, всего две вертикальные: зоны сублитораль и глубинная зона.

Рис. 5.48. Общая схема вертикальной зональности.
Масштаб не соблюден. Изображённые морфоструктурные и гидрологические элементы не были использованы для выделения вертикальных зон, хотя и могут быть их причиной.
Необходимо подчеркнуть недостаточность данных о глубоководной биоте. Разные районы изучены с неодинаковой полнотой, некоторые пока ещё вовсе не обследованы. Большая часть данных относится к сообществам рыхлых грунтов, тогда как сообщества скал практически не изучены. Именно среди последних и были открыты гидротермы.
Краткие характеристики разных вертикальных зон
Верхняя сублитораль
Верхняя сублитораль расположена на глубинах от литорали (включая её) до прибрежного фронта. Глубина, на которой расположен прибрежный фронт зависит от скорости течения (см. выше) и может варьировать от метров до десятков метров. Поскольку скорость течения непостоянна, глубина расположения прибрежного фронта также меняется. В частности, зимой в умеренных и высоких широтах фронт расположен существенно глубже, чем летом. Это обуславливает существование более или менее широкого экотона.
Основные особенности верхней сублиторали, отличающими её от нижней сублиторали приведены ниже, при характеристике этой вертикальной зоны.
— Верхняя сублитораль и воды над ней целиком расположены в прибрежных водах. В холодных и, особенно, умеренных широтах для них свойственна большая амплитуда колебания температуры в течение года. Это приводит к существованию сезонных аспектов: увеличение роли видов с более высокоширотными и/или глубоводными ареалами зимой. В низких широтах они имеют постоянно высокую температуру.
— Непреревное перемешивание всей водной толщи приводит к отсутствию постоянной стратификации (может возникать временная) и обеспечивает содержание биогенов в фотической зоне более высокое, чем в расположенной дальше от берега стратифицированной акватории. Биогены в прибрежные воды поступают также за счёт (1) слабого, но постоянного переноса биогенов из вод открытого шельфа в прибрежные в нижней части термоклина и подстилающих слоях водной толщи (Flint et al., 1996; Поярков, Емельянов, 2001); (2) изменений положения прибрежного фронта: в периоды высоких приливов и штормов фронт смещается глубже, захватывая обогащённые биогенами воды, расположенные ниже пикноклинов; (3) сноса биогенов с суши. В то же время, наличие прикреплённых фототрофов (макрофитов, кораллов и др.) обеспечивает удержание биогенов в прибрежной водной массе. Коралловые рифы могут процветать при содержании биогенов в воде на уровне лимитирующем развитие фитопланктона. И даже в отсутствие макрофитов и кораллов, существенную первичную продукцию на глубинах примерно до 50 м производят донные диатомовые водоросли. В тех случаях, когда интенсивность солнечной радиации на дне > 5% от солнечной радиации на поверхности, продукция донных диатомовых превышает продукцию фитопланктона в данном месте (Longhurst, 2007: 102).
— Сам фронт является местом создания большой первичной продукции, в 1,5—2 раза большей, чем в водных массах, которые он разделяет (Longhurst, 2007). Эта созданная продукция перераспределяется по водным массам, которые фронт разделяет. Поскольку площадь прибрежных водных масс меньше площади стратифицированной акватории, количество принесённой из фронта продукции на единицу площади в них больше.
— Высокая температура и обилие биогенов приводит к высокой первичной продуктивности верхней сублиторали. Её величина мало меняется за вегетационный период, причём первичную продукцию создаёт как фитопланктон, так и бентос (макрофиты, фотосимбиотрофные животные; продукция бентоса может на несколько порядков превышать продукцию фитопланктона). Величина биомассы ассоциаций ламинариевых водорослей обычно 2—25 кг/м2, достигая 660 кг/м2. Поскольку вся эта биомасса создаётся за один год, годовая продукция равна биомассе и достигает, а часто и превышает продукцию наиболее продуктивных наземных сообществ и гораздо выше продукции фитопланктона даже гиперэвтрофных водоёмов (Рис. 5.49). Как указано выше, чистая продукция тропического дождевого леса 0,6—5 кг/м2. Подробнее о сообществах верхней сублиторали см. Жирков (2010).

Рис. 5.49. Сравнительная первичная продуктивность некоторых водных и наземных сообществ.
Водные — горизонтальные линии, пунктир — оценка продукции по биомассе. Наземные (вертикальные линии): 1 — тайга; 2 — Нью-Йорк, сельскохозяйственные зерновые; 3 — широколиственный (неморальный) лес, 4 — Англия, поля сахарной свёклы; 5 — тропический дождевой лес; 6 — поля люцерны, 7 — поля сахарного тростника. По Оdum (1971); Mann (1973); Brinkhuis (1977), Sreenivasan (1972), Бульон (1983), Komiyama et al., 2008; Sargent, Austin (1949, 1954); Odum, Odum (1955); Kohn, Helfrich (1957); Maarel, Franklin (2013), Graham et al., (2017) из Жирков (2010) с добавлениями.
— Высокую первичную продукцию потребляет в основном бентос, в том числе планктонные стадии бентоса. Роль голоплактонных видов незначительна.
Существующие представления о широтной зональности бентали (см. ниже) в значительной степени основаны на данных по этой вертикальной зоне.
Только в верхней сублиторали или преимущественно в ней обитают не только фотосинтетики (макрофиты, включая мангры, герматипные кораллы), но и некоторые крупные таксоны зообентоса не связанные с фотосинтезом, например, плоские морские ежи (обитающие за пределами верхней сублиторали виды по форме тела сходны с неправильными ежами, в отличие от действительно плоских верхнесублиторальных видов), многие семейства рыб, серые киты (Eschrichtius robustus).
Поскольку глубина расположения прибрежного фронта варьирует в широких пределах (от метров до 100 м и более), изучение распределения видов по вертикали в зависимости от глубины существенно размывает границу верхней и нижней сублиторали, делая её значительно менее резкой. Вместе с тем, поскольку на этой границе биота меняется наполовину и даже более, выделение видов, специфичсекой для той или иной зоны не должно быть сложным, причем такие виды должны быть среди массовых. Например у берегов Европы хорошим индикатором верхней сублиторали является Pectinaria koreni, в нижней сублиторали её сменяет P. hyperborea в Белом и Баренцевом моря, а южнее — P. auricoma.
Нижняя сублитораль (= элитораль)
Нижняя сублитораль расположена между прибрежным фронтом и фронтом края шельфа. Фактически весь шельф занят нижней смублиторалью. Для нижней сублиторали характерно разделение воды над ней на поверхностную и подповерхностную водные массы большую часть года: весь вегетационный период в высоких и умеренных широтах и постоянно в низких. Бентос обитает в подповерхностной водной массе, для которой характерна малая амплитуда колебаний температуры. Подповерхностная водная масса находится, за редким исключением, за пределами фотической зоны. Первичная продукция создаётся в поверхностной водной массе, для которой гораздо более низкая первичная продуктивность, причём первичную продукцию создаёт только фитопланктон. Первичная продукция, создаваемая в поверхностной водной массе потребляется в основном в ней же зоопланктоном. Бентос потребляет лишь незначительную её часть, опускающуюся ниже термоклина. В высоких и умеренных широтах поступление детрита на дно имеет весенний (иногда и слабый осенний) пики, соответствующие вспышкам первичной продукции.
Батиаль: верхняя и нижняя
Верхняя батиаль занимает верхнюю часть склона выше главного термоклина, часто непосредственно под фронтом края шельфа. Площадь с глубинами, характерными для верхней батиали (примерно 300—800 м) составляют 3% от площади Мирового океана. Ширина её невелика — обычно 10—20 км. Из-за высокой скорости течения и больших уклонов дна, превышающих крутизну склонов наземных гор, у кромки шельфа развиты жесткие грунты, часто — скалы. Для биоты характерно мощное развитие фильтраторов эпифауны жёстких грунтов, образующих биомассы в несколько кг/м2. В первую очередь это коралловые рифы, образованные во всём Мировом океане всего несколькими видами мадрепоровых кораллов (подробнее см. Жирков, 2010; Келлер, 2011) или губками, высотой в десятки метров и протяженностью в несколько километров. “Эпифиты” этих рифов — брахиоподы, губки, двустворчатые моллюски, усоногие раки, полихеты, мягкие кораллы. Ассоциированная фауна рифов отдельных районов насчитывает сотни и даже более тысячи видов. Одиночные виды склерактиниевых кораллом могут образовывать “луга”, даже на рыхлых (песчаных) грунтах. В районах, где рифы не развиты, их “эпифауна” населяет поверхность грунта (например, биоценозы губок и брахиопод в юго-западной части Баренцева моря). Основными факторами, препятствующими развитию Глубоководных коралловых рифов нет в приполярных районах и в районах с низким уровнем содержания растворённого кислорода (север Индийского и Тихого океанов). По-видимому, эти факторы можно считать основными, определяющими ареал глубоководных коралловых рифов. Из-за сложного рельефа интенсивность придонных течений различается даже в соседних участках, поэтому пятна сестонофагов чередуются с участками с их низкой биомассой. Детритофаги развиты встречаются спорадически из-за редкости подходящих грунтов. Их биомассы невелики (несколько г/м2) также и потому, что в рыхлых грунтах у кромки края шельфа высоко содержание спикул губок и, местами, фрагментов раковин птеропод. И тот и другой компоненты способны существенно повредить ротовые аппараты и пищеварительные тракты, такие грунты и на значительно меньших глубинах очень бедны. Местами на рыхлых грунтах мощные заросли образуют фильтраторы мягких грунтов, в первую очередь морские перья. Количество света, проникающего на глубины до 1 км и более достаточно для охоты, поэтому рыбы верхней батиали имеют крупные глаза. Наличие глаз у хищников и их жертв способствует также широкому развитию биолюминисценции.
Нижняя батиаль расположена под главным термоклином. В отличие от верхней батиали, районы с соответствующими глубинами (обычно >800—1000 м) составляют почти четверть площади Мирового океана (Рис. 5.50). Они расположены не только в нижней части материкового склона, но и на изолированных подводных горах и в системе срединно-океанических хребтов (талассобатиаль), т. е. достаточно равномерно (по сравнению с верхней батиалью) распространены в Мировом океане. Это позволяет биоте этой вертикальной зоны сравнительно легко расселяться по Мировому океану. В нижней батиали получают развитие рыхлые грунты с соответствующей фауной. У подножия, по-видимому, всех материковых склонов, но особенно у дельт крупных рек, имеются выходы метана и других продуктов распада захороненной органики. Уже на глубинах батиали (а в приполярных районах и шельфа) газогидраты стабильны и местами образуют подобные льду кристаллические поля. В основном в пределах батиали расположены и гидротермы. Хотя вклад восстановительных биотопов (гидротерм, сипов, полей газогидратов) в энергию, используемую экосистемами батиали, вряд ли превышает 1%, эту продукцию используют не только обитатели этих биотопов, но и заходящие в них на кормление хищники. Это, а также специфичные для восстановительных биотопов виды существенно увеличивают разнообразие биоты, обитающей в батиали.
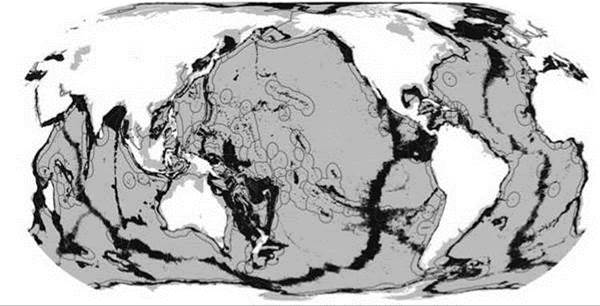
Рис. 5.50. Акватория с глубинами 800—3500 м, темно-серая.
Сплошные тонкие линии — границы национальных экономических зон. Большая часть акватории с глубинами 800—3500 м — подводные хребты и горы. По Watling et al (2013).
Абиссаль и ультраабиссаль
Заселение абиссальных глубин океана многоклеточными происходило как минимум трижды: в докембрии, карбоне-перми и кайнозое. Предшествующая фауна гибла (или, по крайней мере, в значительной степени вымирала) в периоды тёплого климата. Биогеографический ранг ультраабиссали недостаточно установлен, возможно, ультраабиссальные регионы следует рассматривать как отдельные регионы соответствующих абиссальных.
Биота абиссали обитает на значительных глубинах (>2—3 км), где из-за высокого давления, превышающего 200—300 атмосфер, биохимические процессы значительно отличаются от таковых на малых глубинах. Абиссаль расположена глубже точки карбонатной компенсации (см. выше), это существенно увеличивает энергетические затраты на поддержание известкового скелета (иглокожие) или раковин (моллюски), в результате хотя роль иглокожих и моллюсков в абиссали возрастает, роль видов иглокожих с массивными скелетами снижается, а морфология раковин моллюсков упрощается. Иные здесь и доступные ресурсы. Уменьшение же температуры, судя по отчетливой вертикальной зональности полярных регионов, не является фактором, определяющем вертикальную зональность. Группы, дающие в абиссали и ультраабиссали адаптивную радиацию, значительно меняют свою морфологию и поведение. Общая картина отличия биохимии высокого давления от биохимии низкого давления ещё не ясна, пока лишь обнаружены частные особенности. Так, у питающихся грунтом иглокожих (морских звезд и неправильных морских ежей) и обитающих под поверхностью грунта (т. е. в условиях дефицита кислорода), на глубинах шельфа хорошо развиты органы дыхания. У абиссальных же таксонов органы дыхания пародоксальным образом редуцированы (лич. сообщ. А.Н. Миронова). Нектон в абиссали полностью отсутствует, что, по-видимому, связано с малым количеством ресурсов, делающим активное хищничество энергетически невыгодным. В ультраабиссали нет даже быстро движущихся организмов (рыб, головоногих моллюсков, десятиногих ракообразных). Абиссальные рыбы часто лишены глаз (например, Typhlonus nasus, Рис. 5.51). Глубоководные хищники (удильщики Ceratioidei, головоногие моллюски с длинными щупальцами (Mastigoteuthis и др.) используют исключительно стратегию поджидания добычи и не совершают миграций. В абиссали, но не в ультраабиссали широко распространена биолюминесценция.
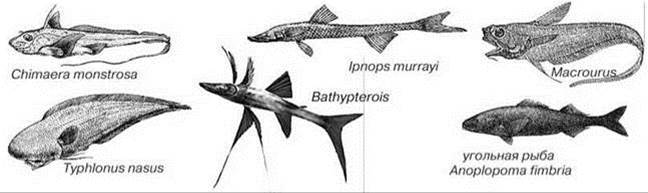
Рис. 5.51. Некоторые глубоководные демерсальные рыбы. По: Парин (1988), Жизнь животных (1971), http://www7a.biglobe.ne.jp/~grafish/fa_u_bathy08.jpg.