Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Шельф - Схемы биогеографического районирования - Биогеография бентали Мирового океана
Как уже отмечено выше при рассмотрении методик, в морской биогеографии исторически сложилось отдельное районирование разных вертикальных зон (и это при том, что сама вертикальная зональность в разных районах океана часто не вписывается в прокрустово ложе сублиторали—батиали—абиссали!). Раздельное рассмотрение разных вертикальных зон возникло в XIX в. и было тогда оправдано малым количеством данных по тому, что тогда называли абиссалью (глубже 100 fms). Поэтому на схеме Ортмана 1896 г. “литораль” (примерно соответствует шельфу в современном понимании) подразделена на ряд областей и подобластей, тогда как подразделения “абиссали” отсутствуют. По мере накопления данных стало ясно, что биота за пределами шельфа тоже неоднородна, но вместо того, чтобы давать единую схему районирования, разные вертикальные зоны продолжали рассматривать раздельно, возможно потому, что эти схемы рисовали разные люди. Число схем “горизонтального” районирования росло с ростом числа выделяемых вертикальных зон. Ряд зоогеографов даже пытается подвести под это теоретическую базу. Так, Е.Ф. Гурьянова обосновывает необходимость отдельного зоогеографического районирования литорали, шельфа, батиали, абиссали и ультраабиссали тем, что “границы донных фаунистических районов, выделенных для разных глубинных зон, не совпадают, а размеры их площадей увеличиваются с переходом от одной зоны к другой, более глубокой” (Гурьянова, 1972: 9). Несовпадение границ в разных вертикальных зонах (вероятно Е.Ф. Гурьянова имела в виду несовпадение границ “по горизонтали”) может быть предметом изучения, но никак не причиной, по которой следует рассматривать тесно связанные между собой биоты по отдельности. Биологического (т. е. единственного, которое следует принимать во внимание при анализе гетерогенности биосферы) обоснования необходимости отдельного районирования разных глубин нет и быть не может, поскольку логическое основание для этого подразделения — глубина — явление вовсе не биологическое. Границы ареалов “по вертикали” принципиально не отличаются от границ ареалов “по горизонтали”. Фактически в случае эврибатных видов глубина, на которой проходит “горизонтальная” граница не является константой, обычно она увеличивается в низких широтах, для этого явления имеется даже специальный термин — субмергенция. Что касается размера площадей фаунистических районов, то применение этого критерия всерьёз обсуждать странно. Неудивительно, что мнение о необходимости раздельного биогеографического районирования разных вертикальных зон не является общепринятым. Так, А.Н. Миронов (1989, 1994) и А.И. Кафанов (1991) считают неправильным отдельное районирование батиали и вышележащих районов и рассматривают их в одной схеме, но, почему-то не разделяя разные вертикальные зоны, несмотря на их существенные фаунистические различия.
Тем не менее, изучение распространения видов по вертикали и горизонтали проводили большей частью отдельно, работ по хорологическому районированию всех вертикальных зон одновременно очень немного, а в масштабах Мирового океана и вовсе нет. Поэтому в настоящее время сложно сопоставить биогеографические регионы, выделенные в пределах хотя бы пяти наиболее часто имеющихся вертикальных зон, тем более, что и в пределах каждой вертикальной зоны нет общепринятой схемы биогеографического районирования. Наименее изучены батиаль и ультраабиссаль, по ним предложено лишь несколько схем. Сравнительно мало данных по абиссали. По биогеографии шельфа данных много, но подходы различных исследователей и применённые ими методы часто различаются весьма сильно, к тому же биогеографическое районирование проводили почти всегда для всего шельфа, не разделяя его на верхнюю сублитораль и элитораль. Несомненно, построение единой схемы биогеографического районирования бентали — дело будущего. Пока же приходится рассматривать частные схемы биогеографического районирования в пределах одной или нескольких вертикальных зон и обсуждать их соответствие друг с другом в Мировом океане. Ниже на примере некоторых акваторий будут рассмотрены конкретные примеры соответствия районирования различных вертикальных зон.
Шельф
Прежде чем перейти к рассмотрению схем биогеографического районирования шельфа, следует напомнить о влиянии плейстоценовых оледенений на биогеографию моря.
1. В время ледниковых максимумов и в периоды межледниковий температура воды в умеренных широтах и высоких широтах существенно отличалась от современных.
2. Во время ледниковых максимумов большая часть современных шельфов была сушей.
Как указано выше, в пределах шельфа проходят существенные биогеографические границы, отделяющие верхнюю и нижнюю сублиторали и биоту кромки шельфа. Однако, хотя схем биогеографического районирования шельфа существует множество, нет схем районирования, учитывающих этот факт. На всех предложенных схемах шельф районируют в целом. Однако, поскольку в первом приближении для обзорных целей можно считать, что фиталь соответствует верхней сублиторали, схемы районирования по макрофитам, герматипным кораллам и другим СС системам с фотосинтетиками — доминантами климакса можно рассматривать как схемы районирования верхней сублиторали, по крайней мере сообществ твёрдых грунтов. Эти частные схемы должны хорошо соответствовать схеме районирования верхней сублиторали в целом.
“Мангры поселяются только в защищённых участках побережий, только на мелководье, т. е. в условиях, которые существовали до их поселения и не были, следовательно, созданы манграми” (Каплин и др., 1991: 446).
Для карты районирования по макрофитам (Рис. 5.59) специфично разделение всей верхней сублиторали на пять областей: две тропических (индо-тихоокеанскую и атлантическую), две умеренных (бореальную и аустральную) и антарктическую.
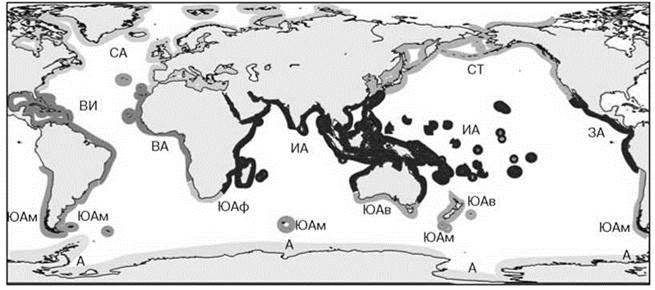
Рис. 5.59. Фитогеографическое районирование Мирового океана по макрофитам Ю.Е. Петрова (1974).
Одной заливкой дана одна область, подобласти обозначены буквами. Бореальная область: СА — Североатлантическая подобласть; СТ — Северотихоокеанская подобласть; Тропико-атлантическая область: ВИ — Вестиндская подобласть; ВА — Восточноатлантическая подобласть; Индо-пацифическая тропическая область: ИА — Индоавстралийская подобласть; ЗА — Западноамериканская подобласть; Аустральная область: ЮАм — Южноамериканская подобласть; ЮАф — Южноафриканская подобласть; ЮАв — Южноавстралийская подобласть; Антарктическая область — А.
Разделение тропического пояса на два региона подтверждают данные по другой группе-эдификатору верхней сублиторали — герматипным кораллам (Рис. 1.16).
На границе тропиков происходит замена доминирующего таксона климаксных ассоциаций. В более высоких широтах на твёрдых субстратах более конкурентоспособными являются СС с климаксом, образованном макрофитами, преимущественно бурыми ламинариевыми водорослями (Рис. 5.60), а в низких — герматипными кораллами (Рис. 5.61) в верхней сублиторали, на литорали — манграми (Рис. 5.62).

Рис. 5.60. Географическое распространение зарослей ламинариевых водорослей и рода климаксных ассоциаций. По Castro, Huber (2003), исправлено.
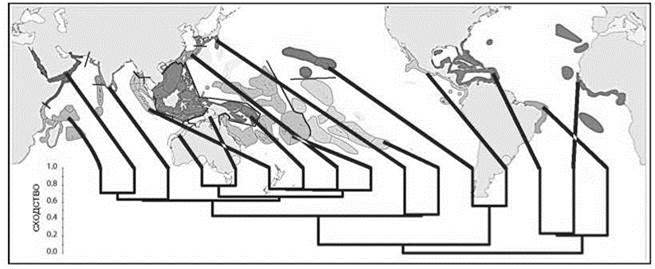
Рис. 5.61. Районирование верхней сублиторали по герматипным кораллам. По Veron et al. (2015).

Рис. 5.62. Ареал мангровых лесов.
На границе тропиков происходит не просто смена таксонов — эдификаторов климаксных ассоциаций. Здесь происходит смена способа фотосинтеза: фотосимбиотрофный в тропиках и фотоавтофный в более высоких широтах. О причинах этого в настоящее время мало что известно. Необходимы исследования как самого симбиоза, так и сравнение биохимии и энергетики фотосинтеза симбиотрофных организмов и макроводорослей, а также биотических взаимоотношений в экосистемах верхней сублиторали. На современном уровне знаний можно отметить следующие моменты:
— В тропиках симбиоавтотрофами являются не только герматипные кораллы и другие Cnidaria, но и моллюски и губки, реже — другие группы. При этом с увеличением широты количество симбиотрофных видов и средняя степень развития симбиотрофии падает. Рассмотрим в качестве примера уже упоминавшихся ранее при анализе Сарматской биоты двустворчатых моллюсков Cardiidae. Это семейство, насчитывающее несколько сотен видов, обладает космополитическими распространением на шельфе, хотя в высоких широтах число видов и невелико (на арктическом шельфе — 2), всё же целое подсемейство Clinocardiinae обитает только в умеренных и холодных водах Северного полушария. Большая часть видов питается, фильтруя придонный слой воды и детрит, лежащий на поверхности грунта. 9 видов специализированных Tridacninae(иногда рассматриваемых как самостоятельное семейство) обитают только в тропиках. Среди них крупнейшие современные двустворчатые моллюски, достигающие размера 137 см и веса более 500 кг (Tridacnagigas). Мантия тридакн содержит большое число симбиотических водорослей. Сама тридакна лежит на дне на коралловых рифах макушкой вниз, развернув и выпятив мантию так, чтобы она занимала максимальную поверхность. 95% синтезированного органического вещества симбиотические водоросли передают хозяину (Flitt, 1993). Продукции симбиотических водорослей, по-видимому, хватает для покрытия по крайней мере большей части энергетических потребностей тридакн, т. е. они занимают нишу фотосинтетиков, хотя все тридакны способны и фильтровать. Это позволяет тридакнам расти со сказочной быстротой — до 10 см в длину в год, при том толщина створок увеличивается на 1 см в год! Детритофагия и симбиоавтотрофия — крайние варианты питания кардид. Есть и виды со смешанным питанием. Даже разные виды Tridacna удовлетворяют свои потребности за счёт симбиотических водорослей в неодинаковой степени. Виды, имеющие симбиотических водорослей, дающих “прибавку” к рациону, но не удовлетворяющих и половины потребностей, есть среди других подсемейств: Fragiinae и даже Clinocardiinae, причём такой симбиоз не столь облигатен. Имеется тенденция снижения числа видов с развитым в той или иной степени симбиозом с водорослями по направлению к полюсам. По-видимому, фототрофное питание с помощью симбиотических водорослей выгодно для вида (энергетически?) только в низких широтах.
— Крупные макрофиты требуют для своего функционирования непрерывного приноса биогенов и уноса продуктов метаболизма. Эти проблемы вовсе не стоят перед герматипными кораллами, поскольку они способны фотосинтезировать при содержании фосфора в окружающей их воде на уровне лимитирования фотосинтеза фитопланктона, а азот симбиотической водоросли поставляют кораллы, усваивающие и продукты обмена водоросли. Заросли крупных бурых макрофитов (саргассы, Turbinaria) в тропиках развиваются только при разрушении коралловых рифов как первая стадия сукцессии. Через несколько лет кораллы вытесняют водоросли. Ламинариевые в тропиках отсутствуют полностью.
— В тропическом поясе обитает много видов, питающихся макрофитами. С продвижением в высокие широты число таких видов резко и быстро падает. Например, рыбы и морские ежи, специализирующиеся на питании морскими травами, отсутствуют в бореальных водах, хотя многочисленны в тропиках (Vermeij, 1978). Устранение видов, питающихся макрофитами в тропиках приводит к бурному развитию водорослей и отмиранию кораллов. Такая смена зарегистрирована, например, в прибрежных водах Вьетнама, где местное население выловило практически всех рыб и крупных беспозвоночных для продажи и питания (личн. сообщ. Д. Астахова). Это показывает, что доминирование кораллов не простое следствие их конкуренции с водорослями, а свойство всей СС.
При более детальном районировании верхней сублиторали следует учитывать, что колебания температура воды в верхней сублиторали за пределами тропиков приводит к появлению сезонных аспектов: в теплые периоды возрастает роль видов с более низкоширотными ареалами, а в холодные — с более высокоширотными.
При проведении действительно биологического районирования Мирового океана следует учесть, что в каждом из минимальных биогеографических выделов (районов) может быть две сукцессионные системы со своими климаксами: сукцессионная система рыхлых грунтов и сукцессионная система твёрдых грунтов. Обоснование этого было дано ранее (Жирков, 2010), здесь же целесообразно остановиться на следствиях.
Прежде всего, надо отметить, что к ассоциациям твёрдых грунтов относится как большая часть ассоциаций макрофитов, так и все мелководные коралловые рифы, т. е. Рис. 5.59, как и Рис. 1.16 — отражают биогеографическе районирование преимущественно ассоциаций твёрдых грунтов.
Рыхлые грунты с точки зрения биоты гораздо менее гетерогенны, нежели твёрдые (скалы, валуны). На них виды могут расходиться, прежде всего, по типу питания (фильтрация придонного слоя воды на разной высоте от грунта, питание поверхностным или подповерхностным детритом, хищничество) и степени углубления в грунт. Часто отмечаемая связь распределения отдельных видов с преобладающим размером фракции грунта (гранулометрией), вполне возможно, является отражением сукцессионной динамики и/или локальных условий местности, в которой проводился анализ, т. е. обусловлена не гранулометрическим составом грунта самим по себе, а не учитываемыми в анализе локальными факторами, связанными с ним. При анализе данных, собранных в гораздо большем районе, локальные факторы теряют свою значимость и результаты получаются совсем иными. Так, мы на материале бентосных съёмок Баренцева моря 1968—1970 гг. проанализировали связь распределения разных видов полихет, питающихся детритом дна, которые, казалось бы должны образовывать более плотные поселения на рыхлых грунтах. Однако анализ, учитывающий влияние одновременно с гранулометрическим составом грунта, глубины и температуры в масштабах всего моря на основании проб с 639 станций показал отсутствие достоверной связи распределения с гранулометрическим составом грунта для многих видов, обитающих и питающихся в толще грунта. В то же время, анализ показал значимую связь абиотических факторов, в частности глубины и гранулометрического состава грунта — на уровне 99,9%.
На твёрдых грунтах возможно расхождение также по неоднородностям субстрата (под камнями, в щелях, на поверхности камней, в норах, вытачиваемых в субстрате и т. д.). Поскольку характер этих условий резко меняется в пространстве, у обитателей твёрдых грунтов гораздо чаще встречаются разные формы территориального поведения. Большая гетерогенность биотопа приводит к тому, что сообщества твёрдых грунтов включают большее число видов, поэтому сравнение суммарного состава биоты биогеографического региона будет в большей степени отражать сходство/различие сукцессионных систем твёрдых грунтов. Виды, входящие в состав сообществ твёрдых грунтов, имеют меньший геологический возраст, нежели виды, входящие в состав сообществ рыхлых грунтов. G.J. Vermeij (1978) проанализировал скорость видообразования в мелководных сообществах по разные стороны Панамского перешейка и обнаружил, что число морфологически сильно разошедшихся видов существенно выше среди видов твёрдых грунтов, чем рыхлых. До образования перешейка этот район был населён единой биотой. Нет оснований полагать, что климатические изменения, происходившие в изучаемых районах, влияли по-разному на обитателей рыхлых и твёрдых грунтов. Поэтому различия в числе морфологически сходных или идентичных видов, обитающих по разные стороны перешейка на разных грунтах, т. е. степень морфологического расхождения исходного вида по разные стороны перешейка, можно считать функцией скорости эволюционных преобразований.
Различия в видовом богатстве сукцессионных систем рыхлых и твёрдых грунтов одного биогеографического региона и различия в скорости их эволюции, возможно, следует учесть в дальнейшем при анализе структуры биосферы.
На Рис. 5.63 — Рис. 5.67 приведены несколько карт биогеографического районирования шельфа Мирового океана. Эти схемы, как и схему на Рис. 5.58 для удобства сравнения я перерисовал, при этом один биогеографический выдел крупного ранга заштрихован одинаково, более крупного ранга — однотипно (например, область заштрихована, подобласти показаны разной штриховкой, другая область залита, подобласти показаны заливкой разной интенсивности и т. п.) и, по возможности, сходно на разных схемах. Более мелкие выделы обозначены буквенными или цифровыми индексами (как в оригинале).
Первая из схем (Рис. 5.62) составлена Е.Ф. Гурьяновой (1964), в ней она попыталась отразить генетические взаимоотношения фаун. Так, например, она объединила в один регион тропические районы восточной Пацифики и западной Атлантики (IV на Рис. 5.63), а также отнесла восточно-атлантический регион от Ла-Манша до о-вов Зелёного мыса, включая Средиземное море, к Западно-Индо-Тихоокеанской области. И то, и другое никем признано не было, поскольку не подтверждается фактическими данными. Особенно это касается отнесения района Средиземного моря и прилежащих районов к Индо-западно-тихоокеанской области. В действительности от биоты Тетиса в Средиземном море ничего не осталось (оно с момента исчезновения Тетиса неоднократно высыхало). Реликтов биоты Тетиса, равно как и их потомков в Средиземном море и прилежащих районах меньше, чем в западной Атлантике. При построении иерархии регионов Е.Ф. Гурьянова использовала метод соответствия ранга регионов рангу эндемичных для них таксонов, который, как показано в Главе 2 при характеристике этого метода, на практике использован быть не может.
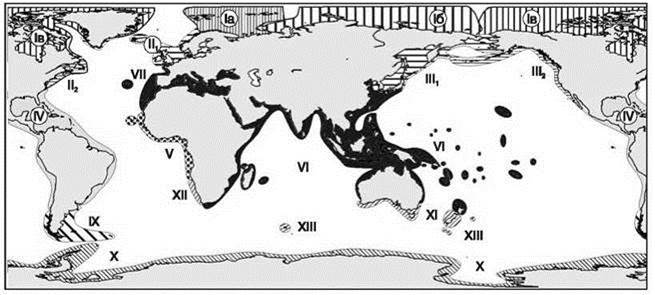
Рис. 5.63. Зоогеографическое районирование шельфа Е.Ф. Гурьяновой (1964).
I — арктическая область: Iа — баренцевоморская провинция; Iб — сибирская провинция; Iв — чукотскоамериканская провинция; II — бореальная северо-атлантическая область: II1 — западно-европейская подобласть; II2 — северо-американская подобласть; III — бореальная северо-тихоокеанская область: III1 — дальневосточная подобласть (беринговоморская провинция; охотоморская провинция; северо-западная япономорская провинция, юго-восточная япономорская провинция, курильская провинция); III2 — орегонская подобласть; IV — тропическая область Центральной и Южной Америки; V — западно-африканская, или гвинейская тропическая область; VI — индийско-западно-тихоокеанская тропическая область (Индо-вест-пацифическая); VII — лузитано-средиземноморская фауна индийско-западно-тихоокеанской области; VIII — северо-чилийская область; IX — антибореальная область; Х — антарктическая область; XI — южно-австралийская, или тасманская область; XII — южно-африканская область; XIII — эндемичные фауны субантарктических островов.
Схема на Рис. 5.64 построена О.Н. Зезиной по брахиоподам. Регионы были выделены методом типизации ареалов и сгруппированы методом оценки сходства списков. К сожалению, в качестве показателя ранга был выбран индекс Престона, который, как показано выше, не следует использовать, в результате ранг выделенных районов фактически не обоснован.

Рис. 5.64. Зоогеографическое районирование шельфа О.Н. Зезиной (1971).
I — бореально-арктическая область: I1 — северо-атлантическая подобласть; I2 — северотихоокеанская подобласть; I2a — азиатско-алеутская провинция; I2б — североамериканская провинция; I3 — арктическая подобласть; II — афмиатлантическая область; II1 — атлантическо-центральноамериканская подобласть: II1a — карибская провинция; II1б — бразильская провинция; II2 — лузитано-мавританская подобласть; II3 — средиземноморская подобласть; III — западноафриканская область; IV — индо-западнотихоокеанская область: IV1 — западноиндоокеанская подобласть; IV2 — индомалайская подобласть; IV3 — японская подобласть; V — тихоокеанско-центральноамериканская область: Va — центрально-американская провинция; Vб — калифорнийская провинция; VI — южнобразильско-урутвайская область; VII — южноафриканская область; VIII — южноавстралийская область; IX — ново-амстердамская область; Х — новозеландско-кертеленская область: Х1 — новозеландская подобласть; X1а — северо-новозеландская провинция, X1б — южно-новозеландская провинция; Х2 — кергеленская подобласть; Х3 — маккуорианская подобласть. XI — чилийский переходный район; XII — антарктическо-южноамериканская область: XII1 — антарктическая подобласть; XII2 — южно-американская подобласть.
Схема на Рис. 5.65 предложена К.Н. Несисом (1982а,б), преимущественно на основе анализа распространения головоногих моллюсков. Регионы выделены методом типизации ареалов, но самостоятельными признаны только регионы, имеющие эндемичные виды (не обязательно головоногих моллюсков). На этой схеме выделено всего три области: одна тропическая и расположенные по обе стороны от неё бореально-арктическая и нотально-антарктическая. Кроме того, выделена одна (? — сам К.Н. Несис пишет об одной, но на его карте показаны три, см. Рис. 5.65) надпровинция переходного характера, а границы между регионами не резкие, а в виде переходных зон. И то, и другое, на мой взгляд, является артефактом использованного подхода к выделению регионов, поскольку фаунистические различия верхней сублиторали и элиторали априорно были признаны биогеографически незначимыми. В действительности в переходной надпровинции Средиземного моря более южные биоты обитают в верхней сублиторали, тогда как более северные — в нижней (элиторали). Совместное рассмотрение в реальности комплементарных друг другу биот создаёт иллюзию единой смешанной (переходной) биоты.
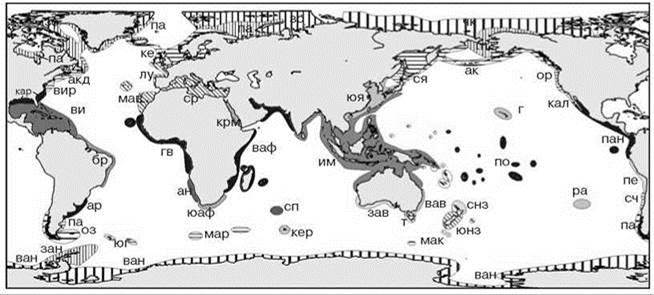
Рис. 5.65. Зоогеографическое районирование шельфа К.Н. Несиса (1982а, б).
Тропическая область с подобластями: западноатлантическая (провинции: кар — каролинская, ви — вестиндская, бр — бразильская, ар — аргентинская), восточно-атлантическая (гв — гвинейская, ан — анголо-намибская, юаф — южноафриканская), индо-вестпацифическая (крм — красноморская, ваф — восточноафриканская, им — индомалайская, юя — южнояпонская (сино-японская), г — гавайская, по — полинезийская, сп — сенпольская, зав — западноавстралийская, вав — восточноавстралийская, снз — североновозеландская, р — рапануйская), восточнотихоокеанская (кал — калифорнийская, пан — панамская, пе — перуанская).
Бореально-арктическая область с подобластями арктической (зс — западносибирская, чк — чукотско-канадская, па — приатлантическая переходная, пт — притихоокеанская переходная), атлантической бореальной (акд — акадийская, ке — кельтская (скандинавская), вир — виргинская), тихоокеанская бореальная (ак — алеутско-камчатская, ся — северояпонская (айнская), ор — орегонская).
Лузитано-средиземноморская надпровинция (л — лузитанская, ср — средиземноморская, мав — мавританская). Нотально-антарктическая область с подобластями: магеллановой (сч — среднечилийская (арауканская) переходная, пат — патагонская, оз — огнеземельская), кергеленской (мар — марионская, кер — кергеленская, мак — маккуорийская), антарктической (юг — южногеоргийская, зан — западноантарктическая (антарктандийская), ван — восточноантарктическая (антарктгондванская).
Тасманийская (т) и южноновозеландская (юнз) провинции не включены ни в одну область, их положение не обсуждается, вероятно, они, аналогично лузитано-средиземноморской, образуют отдельную надпровинцию.
На оригинальной схеме провинциии обычно разделены более или менее широкими переходными зонами, на данной её копии переходные зоны оставлены без картографической нагрузки.
Схема на Рис. 5.66 скомпилирована из отдельных карт монографии J.C. Briggs (1995) и скорректирована по его более поздним работам (Briggs, Bowen, 2012, 2013). Необходимо отметить, что в этих работах в некоторых деталях карты и поясняющие их тексты противоречат друг другу, как в принадлежности провинций к той или иной зоне, так и в числе выделяемых провинций. Приведу лишь три примера: (1) На Fig. 3 (Briggs, Bowen, 2012) показано четыре провинции, а в тексте написано, что их три. (2) В работах Briggs (1995) и Briggs, Bowen (2013) выделена провинция мыса Игольного (Agulhas), она отнесена к субтропической зоне в первой и на Fig. 2 во второй работах и к тропической зоне на Fig. 1 во второй работе. В работе Briggs, Bowen (2012) эта акватория отнесена к субтропическим, но в списке провинций вообще нет субтропических провинций юга Африки (в работе Briggs (1995) их две). (3) Провинция островов Амстердам и Св. Павла перечислена в списке провинций восточно-атлантического региона (Briggs, Bowen, 2012: 13), хотя они находятся в южной части Индийского океана.
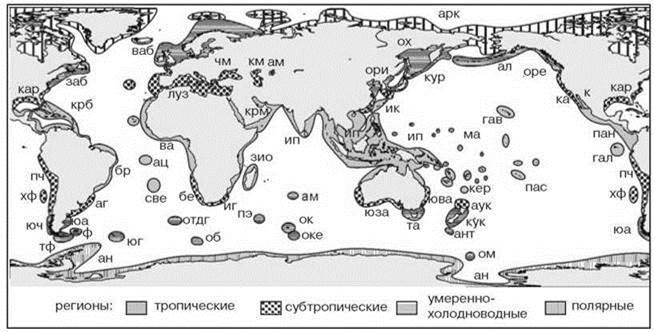
Рис. 5.66. Биогеографическое районирование шельфа Briggs (1995), Briggs, Bowen (2012, 2013).
Тёплые провинции (tropical и warm-temperate): ам — Аральского моря, аук — Ауклендская, ац — острова Ацесьон, бе — Бенгельская, бр — Бразильская, ва — Восточный атлантический, гав — Гавайская, гал — Галапагосская, заб — Западноатлантический бореальный, зио — Западного Индийского океана, иг — мыса Игольного (Agulhas), ип — Индо-полинезийская, к — Кортеса, ка — Калифорнийская (=Сан Диего), кар — Каролинская, кер — Кермадекская, км — Каспийского моря, крб — Карибская, крм — Красного моря, кя — Китаеяпонская, луз — Лузитанская, ма — Маркизская, па — Панамская, пас — острова Пасхи, пч — Перуано-чилийская, све — острова Святой Елены, хф — островов Хуан Фернандес, чм — Черного моря, юва — Восточноавстралийская, юза — Западноавстралийская.
Холодные провинции (cold-temperate и polar): ал — Алеутская, ан — Антарктическая, ант — островов Антиподов, ар — Аргентинская, арк — Арктическая, кур — Курильская, нз — Новозеландская, об — острова Буве, ок — островов Крозе, оке — острова Кергелен, ом — острова Маккуори, оре — Орегонская, ори — Ориентальная, отдг — островов Тристан-да-Кунья и Гоф, ох — Охотоморская, пэ — островов принца Эдварда, та — Тасманский, тф — Терра дель Фуэго, ф — Фольклендская, юа — Южноаргентинская, юг — острова
Южная Георгия, юч — Южночилийская.
“Мы ясно можем видеть, что и Экман, и Бриггс использовали в своих схемах зонально-географический принцип и данные о важнейших климатических границах в морской среде, которым соответствует их границы между областями (“Regions”), и не заметили одну из самых существенных границ у берегов Южной Америки — в районе Огненной Земли и Фолклендских (Мальвинских) островов” (Семёнов, 1982: 215).
Карта J.C. Briggsa отличается более детальной проработкой, чем предыдущие. J.C. Briggs считал отдельной провинцией такую акваторию, которая имеет не менее 10% эндемичных видов (Briggs, 1995: 231). Как показано в Главе 2, критерий доли эндемичных видов можно использовать только при составлении самых первых, прикидочных грубых схем. Обоснованность выделенных районов в отдельных случаях вызывают сомнение, или, наоборот, степень отличия биот не находит адекватного отражения. Briggsвыделял акватории для сравнения, по-видимому, в значительной степени по гидрологическим данным. Это явствует, во-первых, из того, что наиболее крупные регионы его схемы соответствуют регионам, выделенным по температурному режиму, а не по различиям в биоте. Так, он считал равными ранг арктического района и умеренных вод северных Атлантики и Пацифики (Briggs, 1995). Отечественные авторы, более знакомые с этими регионами, или выделяют три равноценных региона для Арктики, умеренной Атлантики и умеренной Пацифики (Е.Ф. Гурьянова) или объединяют арктический регион с каким-либо умеренным (О.Н. Зезина, К.Н. Несис). Впрочем, в последней версии схемы районирования (Briggs, Bowen 2012) ранг холодных и умеренных регионов уравнен. Во-вторых, J.C. Briggs (1995) не проводил биогеографические границы в районах резкой смены биоты, если там нет резких гидрологических градиентов (юг Южной Америки, южная Норвегия). Примечательно, что авторы, перешедшие от метода сравнения списков биот регионов, выделенных по гидрологии к разделению акватории, основанному на непосредственном изучении ареалов видов, пришли к результатам, весьма точно соответствующим результатам В.Н. Семёнова, о необходимости разделения тихоокеанской и атлантической фаун и выделения отдельного региона для южной оконечности Южной Америки (Boschi, Gavio, 2005; Griffiths et al., 2009; Haussermann, Forsterra, 2005; Valdovinos et al., 2003). Эти результаты были признаны Briggs и Bowen (2012, 2013). В основу схем J.C. Briggs положил анализ распространения всех организмов бентоса, но преимущественно рыб, в тропических районах — рыб коралловых рифов. Briggs, Bowen(2012, 2013) использовали также данные по другим рифовым таксоценам. Поскольку видовое разнообразие рифов значительно выше, предлагаемые ими схемы в значительной степени отражают биогеографию верхней сублиторали. В последней версии вместо четырёх зон (тропической, субтропической (warm-temperate), умеренно-холодноводной (cold-temperate) и полярной) выделено всего две зоны, объединившие первые две и последние две зоны прежнего деления. Тропические и субтропические регионыобъединены потому, что “many families and genera span the tropical and warm-temperate regions within each ocean basin, whereas few extend into the cold-temperate regions” (Briggs, Bowen, 2012: 13). Это сходство в значительной степени обусловлено нижнесублиторальными биотами. Как указывает A. Longhurst (1998), температура воды Гвинейского залива (тропический район) на поверхности — 28°С, а на глубине, не превышающей высоту деревьев тропического леса, всего 16°С. При этом видовой состав биоты края шельфа Гвинейского залива весьма сходен с таковым Западной Европы, а температура отличается не более, чем на 2°С, тогда как видовой состав и абиотические условия верхней сублиторали Гвинейского залива и Западной Европы различаются весьма существенно. И здесь, как обычно в биогеографических работах, не вполне адекватные результаты районирования обусловлены изначально методически малоадекватным разделением акватории Мирового океана. Объединение холодных и умеренных регионов также не объяснено (вероятно, причины те же: сходство нижнесублиторальных биот при различии верхнесублиторальных).
Поэтому схема J.C. Briggsa в действительности не чисто бмогеографическая, а эклектичная биогеографо-климатическая.
Схема на Рис. 5.67, преложена Spalding et al., 2007. В этой схеме выделено 232 экорегионов на основе самых разнообразных критериев: ареалам, доминирующим биотопам, геоморфологии, течениям, температуре.
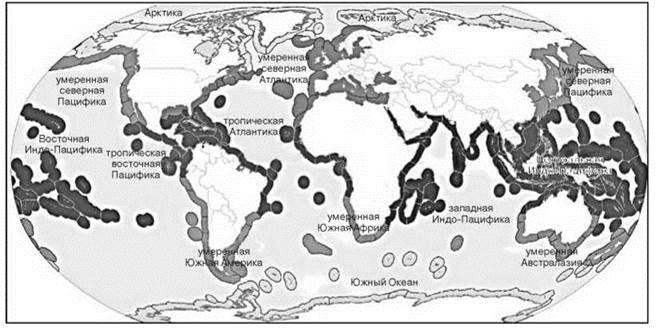
Рис. 5.67 Прибрежные и шельфовые экорегионы. По Spalding et al. (2007).
Деление на провинции и экорегионы дано на цветной вклейке (Рис. Ц12).
Последняя схема (Рис. 5.68) представляет собой составленную мною компиляцию различных схем районирования шельфа. Компиляция была сделана на основе идеологии, описанной в Главах 1 и 2. Несколько замечаний к схеме:
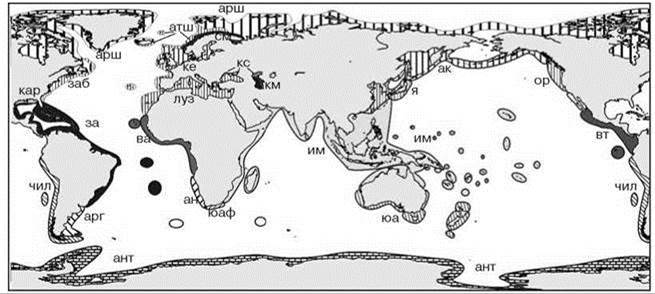
Рис. 5.68. Компилятивная схема биогеографического районирования шельфа.
Аркто-бореальная область с провинциями и/или подобластями: арш — Арктическая шельфовая, заб — Западноатлантическая бореальная, кар — Каролинская, атш — Атлантическая шельфовая, см — Скандинавская мелководная, ке — Кельтская, луз — Лузитанская, я — Японская, ак — Алеутско-камчатская, ор — Орегонская.
Тропическая атлантическая область: за — Западноатлантическая, бр — Бразильская; ва — Восточноатлантическая. Тропическая индо-тихоокеанская область: им — Индо-малайская, вт — Восточнотихоокеанская.
Нотальная (умеренная) область: чил — Чилийская, арг — Аргентинская, ан — Анголо-намибийская, юаф — Южноафриканская, юа — Южноавстралийская, юч — Южночилийская. Антарктическая область: ант — Антарктическая. Каспийская область: км — Каспийская морская, кс — Каспийская солоноватоводная.
— Верхнесублиторальные регионы, несмотря на их сильное отличие от нижнесублиторальных, пришлось, из-за недостатка данных, рассматривать вместе с нижнесублиторальными.
— Схема доведена только до уровня провинций, хотя это и не самое мелкое подразделение. Выделение ареалов ЦС (биогеографических районов) на данной стадии развития биогеографии моря возможно только в районах, где имеется много данных по находкам большого числа видов. Даже для Баренцева моря, акваторию которого изучают более века и в музеях России накоплены обширные коллекции, таких данных недостаточно (Жирков, 2012).
— В одних случаях различные использованные для компиляции схемы давали хорошее совпадение, в других они более или менее существенно различались, особенно в числе районов. При составлении своей компиляции я использовал наименее дробные схемы, поскольку дробность более детальных схем не всегда была на мой взгляд достаточно обоснована.
— В то же время, по-видимому, в большинстве провинций следует разделить шельф на верхнюю сублитораль и элитораль, удалось это сделать только в пределах бореальных районов Северного Ледовитого океана (на арктическом шельфе это разделение, видимо, отсутствует). В любом случае, реальная дробность, конечно, выше, чем приведённая на моей схеме. При этом совершенно не обязательно совпадение границ между районами верхней сублиторали или элиторали. Кроме несовпадения границ верхней и нижней сублиторали, вероятно, как и при переходе от шельфа к батиали и далее к абиссали, будет происходить увеличение размеров биогеографических регионов. Особенно не удовлетворяет меня районирование наших Дальневосточных морей. Однако все предложенные ранее схемы, на мой взгляд, судя по имеющимся у меня данным по распространению полихет и учитывая распространение других групп (двустворчатых моллюсков, иглокожих и др.), также весьма далеки от реальности, в первую очередь из-за совместного рассмотрения всего населения шельфа и, нередко, даже и склона. На данный момент я не в состоянии предложить аргументированную схему районирования, поэтому весь этот район отнесен к одной провинции.
— Районирование Северного Ледовитого океана дано по моим предыдущим работам (Жирков, 2001, 2004, 2010, Jirkov, Leontovich, 2012).
— Районирование Южной Америки дано по В.Н. Семенову (1982) (Рис. 1.58, 2.32).
— В тропическом поясе, основываясь на данных по макрофитам и коралловым рифам, выделено две области: атлантическая и индо-пацифическая.
— Нет разделения бореальных районов у атлантического побережья Северной Америки, традиционно проводимого биогеографами школы ЗИН, здесь я следую американским авторам, лучше знакомым с этим районом (Briggs, 1999; Briggs, Bowen 2012; Briggs, Bowen, 2013), а также биогеографам московской школы.
— Каспийское море рассмотрено как регион ранга области (Зенкевич, 1947; Мордухай-Болтовской, 1960). При этом солоноватоводной биотой, как показано выше, населена только солоноватоводная провинция, к которой отнесены Северный Каспий и распресненные районы Черного и Азовского морей. Средний и Южный Каспий выделены в особую провинцию, населенную морской биотой, сформировавшуюся в условиях моря с пониженной соленостью. Ранее в эту область входило и Аральское море, но в настоящее время его аборигенная биота полностью вымерла и теперь его можно рассматривать скорее, как часть средиземноморского региона.