Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Особенности сукцессионных систем пелагиали - Биогеография пелагиали Мирового океана
При отсутствии градиента абиоты биогеографическая граница в бентали и на суше может существовать как результат взаимодействия разных ценотических систем (ЦС). Неподвижные организмы создают матрицу ЦС с которой взаимодействуют виды, способные передвигаться. На суше матрицу создают растения, в бентали — беспозвоночные и макрофиты. В пелагиали это невозможно, поскольку здесь отсутствие градиента между двумя точками означает, что между ними происходит свободный обмен водой вместе с содержащимися в ней организмами планктона. Это делает невозможным существование и формирование разных ЦС в районах, не разделённых зонами повышенных градиентов. Биогеографические выделы и границы между ними могут появляться только в результате гидрологического обособления вод и водных масс. Зоны резких изменений гидрологических условий соответствуют не только зонам с резкими изменениями абиотических факторов, но и зонам с наименьшим обменом водой, а значит и содержащейся в ней биотой, между районами, которые эта зона разделяет. Поэтому в пелагиали зоны биогеографические границы могут быть только в районах с повышенными градиентами. Конечно, водные массы и круговороты — не замкнутые структуры, непрерывно происходит обмен водой между ними, но интенсивность этого обмена существенно ниже, чем между районами, не разделённым градиентами. Обмен воды приводит к перемещению поперёк биогеографической границы вместе и населения разных биогеографических регионов. Но некоторые водные массы и круговороты замкнуты настолько, что обмен их с окружением не препятствует формированию специфических ЦС.
На неподвижных субстратах (на суше и на морском дне), ЦС, захватив какой-либо участок, способны удерживать его. Кратковременные изменения абиотических условий не влияют на ареалы ЦС благодаря наличию видов, продолжительность жизни которых превышает продолжительность этих изменений. Если при изменении абиотических условий в биогеографическом регионе бентали или суши, сам регион и его биота остаются на месте, то изменение абиотических условий в биогеографическом регионе пелагиали означает изменение системы течений, т. е. изменение положения самого региона. Оледенения вызывали перемещения ЦС бентоса и суши, менялась площадь, которую они занимали, но полностью их биотопы обычно всё же не исчезали, тогда, как последствия перестройки системы течений для пелагиали были гораздо более существенными: многие водные массы исчезали без следа и возникли новые. Современная система течений, особенно умеренных и высоких широт Северного полушария, имеет возраст не более нескольких тысяч лет.
Высокая динамичность границ ареалов биотопов пелагических экосистем делает невозможной даже теоретически существование столь же резких биогеографических границ, как на суше или в бентали. Невозможно и столь же точное проведение биогеографических границ в пелагиали, как это можно сделать в биогеографии бентали, наземной и пресноводной. Подвижность и непрерывность биотопа пелагических биомов приводит и к гораздо меньшей дробности биогеографических подразделений пелагиали, нежели бентали. Это, учитывая весьма высокие расселительные способности многих пелагических видов, приводит к быстрому изменению биогеографической картины вслед за изменением системы течений. Поэтому современная биогеографическая структура пелагиали имеет сравнительно малый возраст. Если в биогеографии пресных вод существенна мезозойская история таксонов, суши — палеогеновая, бентали — неогеновая, то современная биогеографическая картина пелагиали, как указывает К.Н. Несис (1985) имеет плио—плейстоценовый возраст.
Таким образом, в пелагиали, в отличие от суши и бентали, резкость биогеографических границ определяет в первую очередь гидрология, а не взаимодействие ЦС. Размытые гидрологические границы ведут к возникновению обширных участков с промежуточными условиями и смешанным населением — явлению специфичному для пелагиали.
Вся система течений океана гораздо более лабильна, нежели морфология морского дна и условия на нём. Всё это работает против формирования сложных многовидовых комплексов — ЦС, поэтому современные пелагические ЦС, особенно высоких и умеренных широт, в значительной степени имеют признаки группировок. Следствием непрерывно происходящего обмена водных масс и их населения и слабой “эволюционной продвинутости” пелагических ЦС являются большие сложности в таксономии планктона: высокая изменчивость отдельных видов на фоне слабой их морфологической обособленности. Например, таксономией одного из наиболее хорошо известных и массовых рода копепод Calanus, к которому принадлежат наиболее массовые на Земле виды членистоногих, занимались десятки, если не сотни специалистов на протяжении двух с половиной столетий. Однако, несмотря на это, таксономия Calanus, насчитывающего всего 18 видов (WoRMS, 2014), до сих пор окончательно не разработана. Проводимые в последнее время работы по изучению ДНК различных популяций показывают, что, возможно, необходимо дальнейшее дробление видов, которые и без того могут быть сегодня определены только высоко квалифицированными специалистами. Ситуация в других родах гораздо хуже. Таксономические проблемы создают дополнительные трудности в выделении биогеографических регионов на основе ареалов отдельных видов.
Поэтому наиболее простой метод выделения биогеографических районов — метод выделения конкретных биот (см. Главу 2) — даёт не столь удовлетворительные результаты, как на суше и в бентали. Дополнительная трудность, с которой сталкиваются биогеографы: любой участок поверхности моря выглядит очень похожим на все остальные. Вместе с тем, поскольку биогеографические регионы определяет гидрология, существует довольно обоснованная тенденция использовать непосредственно гидрологические данные для биогеографического районирования. Однако их непосредственное использование гидрологических данных затрудняет существенное их изменение во времени (например, осцилляции Эль Ниньо), поэтому биогеографические границы, проведённые на основе многолетних данных, значительно менее резки, чем в каждый конкретный момент, т. е. реально существующие.
Малая продолжительность жизненных циклов фитопланктона и отсутствие почвы или её аналогов позволяет пренебречь сукцессиями, идущими более одного года (в отличие от биогеографии суши и бентоса) и проводить непосредственный анализ функционирования пелагических экосистем. Такой анализ позволяет выделить разные типы экосистем поверхностных вод (ЦС), ареалами которых и являются биогеографические регионы.
В функционировании пелагических ЦС существенную роль играет микробная петля (microbial loop) — утилизация растворённого органического вещества бактериопланктоном, который делает его доступным для более высоких трофических уровней (Рис. 6.6 и 6.7).
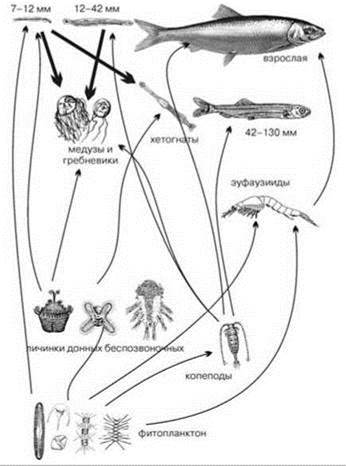
Рис. 6.6. Классическая схема пищевых цепей пелагиали (в качестве нектона показана сельдь на разных стадиях развития).
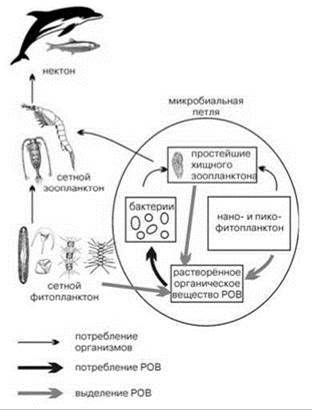
Рис. 6.7. Схема пищевых цепей пелагиали, учитывающая микробную петлю.
Количество световой энергии, необходимой для фотосинтеза падает с глубиной. На некоторой глубине энергия, получаемая с помощью фотосинтеза, становится равной энергии, которую тратит организм на дыхание. Такую глубину называют компенсационная глубина. Фитопланктон, в отличие от зоопланктона не способен к вертикальным миграциям, поэтому ему необходимо находиться выше компенсационной точки. Для уменьшения скорости погружения фотосинтетики пелагиали имеют разнообразные структуры, обычно это выросты клеток. Всё же, несмотря на наличие таких структур, они погружаются и, если гидрологическая структура не препятствует их погружению ниже компенсационной глубины (отсутствует пикноклин), убыль клеток может стать равной или даже превысить их образование в результате размножения, а величина первичной продукции стать малой или даже отрицательной. Во многих районах величина первичной продукции значительно и закономерно варьирует в течение года. Периоды повешенной продуктивности называют вспышками или цветением фитопланктона. За редким исключением непременным условием возникновения вспышек как в море, так и в пресных водах является возникновение пикноклина, расположенного на глубинах меньших глубины компенсационной точки. Природа пикноклина в разных районах различна. Пикноклином может быть или термоклин и/или галоклин. Влияние солёности на плотность гораздо выше, чем температуры (см. Главу 5), поэтому, если пикноклин — это одновременно и галоклин и термоклин, влиянием температуры на плотность можно пренебречь. При галоклине солёность на поверхности ниже, чем под галоклином, при термоклине температура воды у поверхности обычно выше, чем под термоклином. Поскольку зависимость плотности от температуры не монотонная, в воде с солёностью меньше 24,695%о при температуре близкой к точке замерзания её плотность меньше, чем при более высокой. В наиболее распространённом случае — в пресной воде — вода имеет максимальную плотность при 4°С, и меньше при более низкой (Рис. 5.18). В таких случаях температуру воды над термоклином может ниже, чем под ним, способствуя возникновению цветения. Такое наблюдается, например, в Байкале (см. Главу 4), где весеннее цветение происходит подо льдом. В некоторых районах вспышки продукции могут происходить даже при отсутствии стабилизации поверхностного слоя. В этих случаях их обеспечивает пикопланктон, обладающей скоростью размножения достаточно высокой, чтобы компенсировать погружиющиеся клетки.
Фотосинтезирующие организмы нуждаются в биогенах. Основные биогены в океане — азот и фосфор (входящий на суше в состав биогенов калий в море есть в избытке), в отдельных случаях и необходимы и другие элементы, например для фиксации азота атмосферы необходимо железо. Чем меньше размер клеток фитопланктона, тем меньшие максимальные биомассы они создают в природе*, и тем меньшая концентрация минерального азота лимитирует их рост. Поэтому рост биомассы фитопланктона во время сезонных вспышек, или цветения (bloom), там, где их наблюдают, обеспечивают водоросли с крупными клетками (диатомовые и динофлагелляты), а в бедных биогенами водах основную продукцию создают виды с мелкими клетками, независимо от того, постоянен дефицит или имеет сезонный характер.
Динамика продуктивности различна в разных районах: она может быть примерно постоянной или значительно варьировать по разным причинам. В соответствии с динамикой продуктивности в течение года и с учетом причин, вызывающих её можно выделить шесть основных типов динамики первичной продукции (Longhurst, 2007).
1. Единственный период цветения во время увеличения освещённости весной. Характерна для полярных биомов.
2. Весеннее сильно выраженное цветение, возникающее обычно после установления сезонного пикноклина (классическая динамика Сведрупа). Цветение заканчивается после исчерпания биогенов и подъёма фитофагов с глубины. Осенью, после опускания фитофагов на глубину, может происходить второй, гораздо более слабый пик после исчезновения пикноклина и поступления биогенов на глубины меньшие компенсационной. Характерна для биомов западных ветров.
3. Зимне-весеннее слабо выраженное цветение. Кроме незначительной амплитуды и низкого уровня первичной продукции отличается от предыдущего варианта более длительным периодом цветения и тем, что оно заканчивается в основом из-за исчерпания биогенов, роль фитофагов при этом незначительна.
4. Постоянно низкий уровень первичной продукции и хорошая сбалансированность продукции и потребления. Компенсационная глубина расположена ниже пикноклина, поэтому максимум концентрации хлорофилла лежит в пределах пикнолина. Характерна для биома пассатов.
5. Зависящая от муссонов динамика. Этот тип характерен для тропических районов с низкой первичной продукцией где направление пассатов меняется два раза в год (муссоны). В начале каждого периода муссонов происходит заглубление пикноклина ниже компенсационной глубины. Это приводит к появлению четырёх сезонов в динамике первичной продукции.
6. Динамику первичной продукции определяет интенсивность апвеллингов. Как и в биомах западных ветров фитофаги совершают вертикальные онтогенетические миграции. Поднимающаяся в апвеллингах вода, таким образом, приносит не только биогены, но и фитофагов.
Акватории, для которых характерны эти шесть типов динамики первичной продукции показаны на Рис. 6.8.
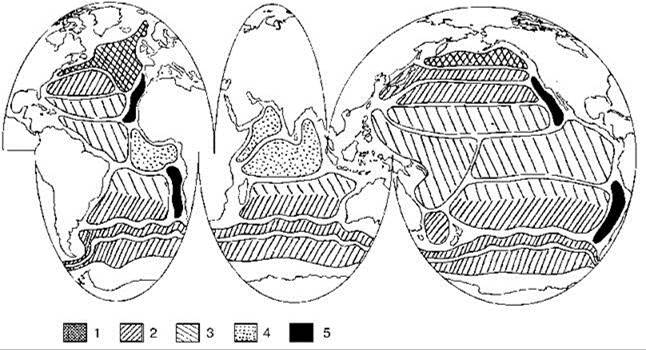
Рис. 6.8. Распространение разных типов сезонного цикла продукции фитопланктона. По Longhurst (2001).
1 — биомы западных ветров, весеннее цветение, лимитированное биогенами, 2 — биомы западных ветров, зимне-весеннее цветение, лимитированное биогенами, 3 — биомы пассатов, сезонная изменчивость продукции невелика, 4 — биомы пассатов, большая изменчивость продуктивности из-за муссонов, 5 — неритические биомы апвеллингов.
* Не в лаборатории!