Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Стратегии жизненных циклов - Ценофильные и ценофобные виды - Сукцессионная система - Био-экология, или синэкология
Стремление обобщить накопленную информацию по аутоэкологии видов привело к возникновению концепций стратегий жизненных циклов (или просто жизненных стратегий), называемых также эколого-ценотическими стратегиями. В конечном итоге, во всех концепциях рассматривают распределение энергии между различными популяционными процессами (миграции, особенности размножения) и особенностями биологии отдельных особей (рост, размеры, продолжительность жизни и т. п.). Различные концепции рассматривают распределение аутэкологических характеристик по различным осям, которые авторы данных концепций считают наиболее значимыми.
Классификация, рассматривающая две жизненные стратегии — r-К
Самая простая классификация жизненных стратегий выделяет два основных типа. Названия этих типов различны. В первом варианте (McLeod, 1884, цит. по: Миркин и др., 2002) их определили, как “капиталисты” и “пролетарии”, сейчас их называют K- и r-стратегами (MacArthur, Wilson, 1967; Пианка, 1981; Ильяш и др., 2003) по коэффициентам логистического уравнения роста численности популяции r и К (Рис. 1.43). Согласно одной из теорий (весьма популярной в современной экологии) и отдельным подтверждающим её примерам, в сообществах с большим числом видов конкуренция — основной фактор отбора в условиях дефицита ресурсов, дефицит ресурсов же считают непременным свойством природных сообществ. Иными словами, конкуренцию за ресурс, согласно таким представлениям, считают основным фактором взаимодействия видов. Для многих экосистем или гильдий (например, сосудистых растений в наземных экосистемах или фитопланктона) это вполне допустимое упрощение. Анализируя логистическую модель роста численности популяции, MacArthur и Wilson (1967) предположили, что в стабильных условиях конкуренция за ресурс приводит к отбору видов, у которых основной поток энергии через популяцию направлен на продуцирование более конкурентоспособных потомков (К-стратегия). Но большие затраты на производство отдельного потомка приводят к ограничению числа потомков, продуцируемых половозрелой особью. Это ведет к снижению максимальной скорости роста и снижению расселительных способностей вида. В экосистемах, где численность популяции ограничивают не лимитирующие факторы, за которые идет конкурентная борьба, а внешние воздействия (пресс хищников, постоянно действующие нарушения и т. п.), преимущество получают виды с более высокой скоростью размножения (r-стратеги). Эти виды производят большое количество потомков, на производство каждого из которых затрачена гораздо меньшая энергия. Напряжённость межвидовой конкуренции и связанная с ней степень доступности лимитирующего ресурса определяют положение видов на оси r — и К-стратегий. К-виды направляют основной поток энергии на поддержание жизнедеятельности взрослых особей, при r-виды — на размножение.
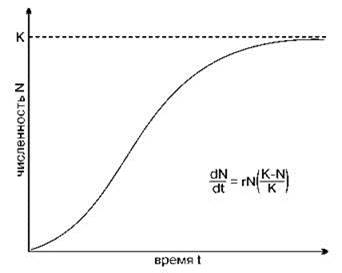
Рис. 1.43. Логистическая кривая роста численности популяции.
К — предельная численность вида, r — константа, отражающая максимально возможную скорость роста популяции.
Э. Пианка (1970, 1981) предположил, что в слабо нарушаемых сообществах преобладают К-виды, а в сильно нарушаемых — r-виды. С биологической точки зрения очень важна причина нарушений: являются ли они естественными, свойственными самим экосистемам, или их биотопам; или же это антропогенные воздействия. К естественным нарушениям, в отличие от антропогенных, виды эволюционно адаптированы. Однако, этот аспект теория Э. Пианка не затрагивает.
R- и К-стратегии — крайние варианты, в реальных же условиях наблюдается континуум переходных форм.
На практике для определения положения видов на оси r-К используют различные показатели. Часто указывают, что для r-стратегов (их называют также оппортунистами) характерен короткий жизненный цикл с единственным периодом размножения в раннем возрасте и большой расход энергии на репродуктивные усилия. Их считают характерными для эфемерных биотопов. Наоборот, для К-стратегов считают характерной долгую жизнь, многократное размножение, начинающееся сравнительно поздно. Но при изучении характеристик реальных видов эти показатели нередко дают противоречивую картину. Так, для подавляющего большинства видов морского макробентоса, а также многих рыб и головоногих, на суше — для деревьев, характерны высокая плодовитость, большой расход энергии на репродуктивные усилия, нередко сочетающиеся с большой продолжительностью жизни, причём плодовитость растёт с возрастом и размером. Более того, известны случаи, когда разные полы одного вида придерживаются различных стратегий или даже разные особи одного пола придерживаются неодинаковых стратегий размножения. Это показано, например, для головоногих моллюсков (Лаптиховский, 2006). Поэтому при попытках отнесения к r-К-стратегам не отдельных специально подобранных видов, а достаточно полного набора видов из одной экосистемы обычно приходится говорить о свойствах r-К-статегов, при этом большую часть видов часто не удаётся отнести ни к тем, ни к другим. Тем не менее, r-К-классификация пользуется большой популярностью, в первую очередь в педагогике из-за кажущейся её простоты и логичности. Сомнения в её адекватности возникают только при переходе от рассмотрения идеальных ситуаций к реальным.
Классификации, рассматривающие три жизненные стратегии
Недостатки r-К-классификации пытаются устранить в других классификациях. Широкое распространение получили классификации жизненных стратегий видов, выделяющие три основных типа стратегий. Такие классификации обычно имеют не одно, а два логических основания. Первым такую классификацию предложил геоботаник Л.Г. Раменский (1938). Он назвал виды с тремя основными типами стратегий:
— Виоленты (от латинского violentus — слишком сильный), или “виды-львы”, “виды-силовики” — “энергично развиваясь, они захватывают территорию и удерживают её за собой, подавляя, заглушая соперников энергией жизнедеятельности и полнотой использования ресурсов среды” (Раменский, 1938: 279).
— Патиенты (от латинского patientis — выносливый, терпеливый, способный переносить), или “виды-выносливцы” — “уподобляются верблюду... в борьбе за существование они берут не энергией жизнедеятельности и роста, а своей выносливостью к крайним, суровым условиям, постоянным или временным... В мезофильных местообитаниях они нередко прекрасно развиваются, но в природе заглушаются растениями первой группы (виолентами). Патиенты достигают господства в условиях крайних и для них же часто менее благоприятных, притом иногда чрезвычайно различных” (Раменский, 1938: 279-280).
— Эксплеренты (от латинского expletum — наполнять, заполнять), или выполняющие — “фигурально уподобляющиеся шакалам, имеют очень низкую конкурентную мощность, но зато они способны очень быстро захватывать освобождающиеся территории, выполняя промежутки между более сильными растениями; так же легко они и вытесняются последними” (Раменский, 1938: 280).
“Отечественный ученый Л.Г Раменский предложил классифицировать предприятия и компании по четырем типам инновационного поведения: предприятия-виоленты (виолентное поведение); предприятия-патиенты (патиентное поведение); предприятия-эксплеренты (эксплерентное поведение); предприятия-коммутанты (коммутантное поведение). Аналогичная классификация предложена швейцарским экономистом Х.Фризевинкелем, который проводит аналогию инновационного поведения компаний с поведением в животном мире: виоленты — львы, слоны, бегемоты; патиенты — лисы; эксплеренты — ласточки; коммутанты — мыши” (Новиков, 2007: 53).
Хотя в своей классификации Л.Г. Раменский прямо не выделял её логические основания, из приведённых цитат ясно, что это конкурентоспособность и устойчивость к неблагоприятным условиям. Независимо от Л.Г. Раменского аналогичную классификацию предложил L. Van Valen (1971), идеи которого были развиты англичанином J.P. Grime(1974, 1977, 1979, 1982) применительно к наземным сосудистым растениям и американцем G.J. Vermeij (1978) применительно к морскому бентосу. Они предложили другие термины: competitor, или biotically compitent species (биотически успешные виды), stress tolerant (виды, устойчивые к стрессу) и ruderal (рудеральные, сорные виды), поэтому данную классификацию называют по первым буквам C-S-R.
J.P. Grime в качестве логических оснований в этой классификации использовал относительную скорость роста особи и морфологический индекс (развитие надземной части растения) (Табл. 1.7, Рис. 1.44). Сопоставляя свою классификацию с r-K, J.P. Grime считал, что C-виды = К-видам, R-виды = r-видам, а S-виды не имеют аналогов в r-Kклассификации.
Табл. 1.7. C-S-R-стратегии по J.P. Grime.
|
конкурентоспособность, |
скорость роста |
|
|
размеры |
высокая |
низкая |
|
высокая, крупные низкая, мелкие |
С — виоленты: высокая скорость роста, крупные размеры R — эксплеренты или рудералы: мелкие, высекая скорость роста + “банк семян” для переживания нарушений |
S — патиенты или стресс-толе- ранты: низкая скорость роста, небольшие размеры |

Рис. 1.44. Треугольник Грайма.
C, R, S — основные типы жизненных стратегий, CR, CS, RS, CRS — переходные между ними типы жизненных стратегий. По Миркин и др. (2001).
Б.М. Миркин с соавторами (2002), характеризуя три типа стратегий, относит к виолентам “мощные растения”, произрастающие в отсутствии нарушений. Это обычно — деревья, кустарники или высокие травы. Они “держат под контролем условия среды и полностью (или почти полностью) используют обильные ресурсы таких местообитаний” (с. 42—43). Такие растения конкурентно подавляют остальные, но происходит это только при наличии значительных ресурсов. Авторы приводят пример с канареечником (Phalaroides arundinacea), виолентность которого можно обнаружить только при внесении большого количества минеральных удобрений. Фактически все виоленты, в их понимании — резко выраженные эдификаторы, а также, по-видимому, любые виды, способные образовывать густые одновидовые поселения, т. е. логическим основанием для их выделения являются не внешние условия, как у Л.Г. Раменского, а морфология взрослого растения, как у J.P. Grime, причём в отличие от последнего, конкурентоспособность видов Б.М. Миркин с соавторами вовсе во внимание не принимают. При ухудшении условий конкурентное преимущество получают виды, способные их переносить — патиенты. Среди патиентов авторы выделяют экотопические и фитоценотические. Экотопические патиенты произрастают в местах, где условия плохи сами по себе, а фитоценотические — в местах, где плохие условия создают эдификаторы-виоленты. В местах, где нарушения ухудшают условия, растительный покров образуют эксплеренты. Это однолетние (реже малолетние) растения с большой семенной продукцией. Их семена попадают в недавно нарушенные места благодаря высоким рассели- тельным способностям, или же они уже были в почве в этом месте, дожидаясь подходящих условий. Кроме настоящих эксплерентов Б.М. Миркин с соавторами (2002) выделяет ложных эксплерентов, которые постоянно входят в состав растительных сообществ, образованных виолентами или патиентами, но вегетируют в короткие периоды, когда имеющиеся ресурсы почему-либо недоступны доминантам. Сюда авторы относят растения, которые обычно называют эфемероидами и эфемерами: вегетирующие в листопадных лесах до распускания листвы, в пустынях в периоды дождей.
G.J. Vermeij (1978) считает, что логическими основаниями для выделения трёх типов стратегий являются особенности окружающей среды (судя по тексту — благоприятность условий, в частности, их стабильность) и то, как виды реагируют на них. Он полагает, что для видов с высокой скоростью роста популяции, которых он называет не эксплеренты, а оппортунисты (т. е. считает их синонимом r-стратегов), характерна высокая способность к колонизации эфемерных или часто нарушаемых биотопов благодаря высокой способности к расселению. Однако их конкурентоспособность низка, поэтому виды других групп вытесняют их. Для патиентов характерна способность длительно существовать в неблагоприятных условиях (выносить физиологический стресс). Их отличает низкая скорость роста и, независимо от условий их существования, невысокий репродуктивный потенциал. В результате при благоприятных условиях, они также растут очень медленно, поэтому их конкурентоспособность низка. В качестве примера таких видов он приводит виды супралиторали и верхних горизонтов литорали, абиссали и районов с пониженной солёностью. Многие черты К-стратегов возможно связаны не с их высокой конкурентоспособностью, а с устойчивостью особей таких видов, поэтому К-стратеги и патиенты могут иметь немало сходного в своей биологии. К третьей группе он относит биотически успешные виды, обитающие в условиях, близких к оптимальным. В эту группу попадают не только К-стратеги. Если К-стратеги по определению поддерживают плотность своих популяций на высоком уровне благодаря конкурентному превосходству во взаимодействии с другими видами, то биотическая успешность видов, по мнению G.J. Vermeij, может иметь и другие причины. Биотически успешные виды также имеют большую продолжительность жизни, сравнительно высокую скорость роста на ранних стадиях развития. В море, по его мнению, такие виды преобладают на наружной поверхности рифов, в верхних нескольких сантиметрах рыхлых грунтов мелководий, в нижней литорали.
Многие живые организмы имеют короткую продолжительность жизни и не имеют выраженного периода индивидуального роста или, по крайней мере, не имеют выраженной возрастной структуры популяции (простейшие, фитопланктон, многие виды зоопланктона, особенно пресноводного и др.), поэтому к ним не применимы логические основания классификаций, использованные Л.Г. Раменским, L. Van Valen, J.P. Grime и G.J. Vermeij.
Ю.Э. Романовский (1989а,б), изучавший пресноводный зоопланктон, обратил внимание на то, что в природе обычно ресурсы испытывают флуктуации и в качестве логического основания предложил использовать амплитуду и частоту колебания ресурса. По его мнению, при больших амплитудах колебания ресурса высокой численности популяции достигают r-стратеги (ярко выраженные эксплеренты), способные быстро увеличить численность. При низкой амплитуде высокой численности популяции достигают К-стратеги (крайние патиенты), которые не только сами устойчивы к минимальной концентрации ресурса, но и “поддерживают его количество на постоянном и низком уровне, недоступном для других видов” (1989а: 25). С другой стороны, продолжительность периодов дефицита ресурса также различна. Если период дефицита ресурса существенно превышает продолжительности жизни особей, то такие r-стратеги (эксплеренты) переживают этот период на латентной стадии (споры и семена некоторых растений, покоящиеся яйца беспозвоночных, а также особи, способные впадать в состояние анабиоза). Если период дефицита ресурса меньше продолжительности жизни особей, то такие r-стратеги (виоленты) переживают его на половозрелой стадии. Таким образом, С-S-R-стратегии — это адаптации к разным уровням колебания ресурсов независимо от природы таких колебаний, и положение вида в С-S-R-пространстве определяет распределение расхода энергии на различных стадиях онтогенеза (Рис. 1.45).
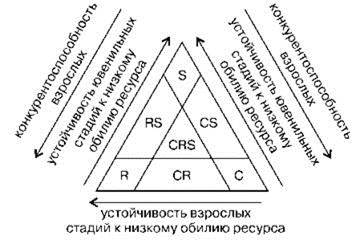
Рис. 1.45. C-S-R-стратегии у зоопланктона. По Ю.Э. Романвскому (1989).
Л.В. Ильяш (Ильяш и др., 2003) на основе анализа популяционных стратегий фитопланктона в качестве логических оснований классификации стратегий предложила использовать способность вида конкурентно воздействовать на совместно обитающие с ним виды и чувствительность к такому воздействию со стороны других видов. При увеличении значений одного показателя уменьшается другой и наоборот. Эксплеренты (R-стратегия) и виоленты (С-стратегия) занимают при этом крайние положения, патиенты (S-стратегия) — среднее, одновременно только для них характерна высокая устойчивость к низкой концентрации ресурса (Рис. 1.46).
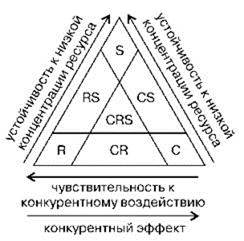
Рис. 1.46. C-S-R-стратегии у фитопланктона. По Л.В. Ильяш и др. (2003).
Сравнивая классификации разных авторов, можно отметить, что все выделяют три типа стратегий:
— Оппортунисты, или эксплеренты — виды, которые быстрее остальных начинают потреблять новый ресурс (территорию, биогены и т. п). Способы реализации этой стратегии весьма разнообразны. Одни виды имеют специальные стадии для пережидания периодов, когда ресурс малодоступен. Другие, благодаря относительно короткому жизненному циклу (способности быстро увеличить свою численность) и высокой расселительной способности, дают вспышки численности в местах, где ресурс временно становится доступным. Особи третьих сами мигрируют между местами с доступным ресурсом, потенциал размножения и расселения при этом могут быть и невысокими. Примером вида с такой стратегией Л.Г. Раменский считал шакала.
— Патиенты — виды, особи которых обитают в неблагоприятных условиях. Существует слишком большая субъективность в отнесении условий к благоприятным или нет и, соответственно, отнесения видов к виолентам или патиентам. Так, условия жизни в абиссали или в засушливых районах считают неблагоприятными. Виды, которые там существуют относят к патиентам, поскольку они способны переносить эти неблагоприятные условия. Вместе с тем, наличие в таких условиях процветающих популяций патиентов позволяет интерпретировать данные условия как благоприятные для них. Правильнее было бы оценивать качество биотопа с точки зрения самого вида, а не наших представлениях о том, что для него хорошо, а что плохо. Условия в пустыне неблагоприятны для зостеры, но благоприятны для кактуса, и наоборот, условия верхней сублиторали неблагоприятны для кактуса и благоприятны для зостеры. А для солероса (Salicornia europaea), растущего и там, и там, благоприятными, по-видимому, следует считать оба биотопа. Отсутствие каких-либо объективных критериев благоприятности-неблагоприятности видно из следующего примера. Л.Г. Раменский (1938), иллюстрируя жизненную стратегию патиентов, приводит всего два примера: сосну (Pinus sylvestis) и тростник (Phragmites australis). Характеризуя последний, он пишет: “Тростник — прямо специалист по разнообразным трудным условиям, настоящий земноводный верблюд” (стр. 280). Б.М. Миркин с соавторами (2002) приводит два примера виолентов (sic!): канареечника и того же тростника (Миркин и др., 2002: 42). Представленный выше пример канареечника также хорошо иллюстрирует субъективность критериев. “Виолентность” вида проявляется только при внесении минеральных удобрений, т. е. только при антропогенном воздействии. Следуя этой логике, к ярко выраженным виолентам следует отнести и все остальные культурные и декоративные растения, “виолентность” которых проявляется при других воздействиях человека (обработка гербицидами, регулярная вспашка и прополка и т. п.). С точки зрения Л.Г. Раменского, канареечник следовало бы считать типичным эксплерентом, поскольку ни о какой “высокой энергии жизнедеятельности” в его случае не приходится говорить, ибо “на контрольном участке, где удобрения не вносились... к шестому году... канареечник был практически полностью вытеснен” (Миркин и др., 2002: 43). С другой стороны, латентные стадии оппортунистов не всегда легко (а то и невозможно) отличить от особей патиентов.
— Все остальные виды относят к третьей группе, которую различные авторы называют по-разному. Наиболее нейтральным (а, следовательно, более универсальным) является термин биотически успешные виды.
“Эти добрые люди, — заговорил арестант и, торопливо прибавив: — игемон, — продолжал: — ничему не учились и все перепутали, что я говорил. Я вообще начинаю опасаться, что путаница эта будет продолжаться очень долгое время. И все из-за того, что он неверно записывает за мной” (Булгаков, 1967. “Мастер и Маргарита”).
При всём внешнем сходстве разные классификации, выделяющие три жизненные стратегии, неодинаковы, поскольку при их проведении данные авторы использовали различные логические основания. Причина этого состоит в том, что из множества вероятных логических оснований авторы классификаций выбирают те, которые считают наиболее существенными. Представление же о существенном неодинаково у различных исследователей, во-первых, потому, что на разных стадиях развития науки эти представления меняются. Во-вторых, каждый автор строит свою классификацию для изучаемых им групп: Л.Г. Раменский, L. Van Valen, J.P. Grime и Б.М. Миркин с соавторами — для наземных высших растений, G.J. Vermeij — для макробентоса, Ю.Э. Романовский — для пресноводного зоопланктона, Л.В. Ильяш — для фитопланктона. Очевидное несходство биологии данных групп приводит к существенным различиям в возможностях реализации жизненных стратегий. К тому же эти группы входят в состав разных экосистем, а особенности экосистем и биологии доминирующих в них групп в значительной степени взаимосвязаны. Например, на коралловом рифе или в лесу очень важна эдификаторная роль отдельных видов, тогда как в фитопланктоне этот эффект гораздо менее значим. Всё это делает неизбежным существование многочисленных классификаций жизненных стратегий, что мы и наблюдаем в реальности. Созданные каждая для конкретной ситуации, различные классификации хорошо их описывают. В то же время, одновременное существование разных классификаций делает каждую менее универсальной, а применимость конкретных классификаций в экосистемах другого типа часто сомнительна. Субъективность выбора логического основания для классификации может сопровождаться субъективностью оценки положения вида на градиенте факторов, по которым проводится классификация. Как показано выше на примере благоприятности-неблагоприятности, одни и те же условия (для одного и того же вида!) разные авторы могут трактовать диаметрально противоположно; это резко обесценивает такие классификации и снижает их общетеоретическую ценность.
Как и в случае r-К-стратегий, три стратегии рассматривают идеальные случаи. Реально же виды образуют континуум в С-S-R-пространстве. Также, как и в случае r-К-стратегий, подразделение видов, удобно с педагогической точки зрения (хотя, как показывает практика, и менее, чем r-К-стратегии). На практике в отношении каждого вида выполнимо с теми или иными оговорками.
Большое внимание, уделяемое обсуждению различных жизненных стратегий, связано со стремлением через жизненные стратегии видов объяснить поведение экосистем. Такое возможно только если в экосистемах преобладают неспецифические взаимодействия видов. Накопленные экологией данные свидетельствуют о том, что экосистемам присущи особенности, характерные только для них. Эти особенности возникают лишь в результате взаимодействия её частей (видов), специфических для каждой экосистемы. Такие особенности не могут быть отражены в классификациях жизненных стратегий видов. Например, во многих экосистемах велика роль “ключевых видов”, но это — характеристика роли вида в экосистеме, а не его жизненной стратегии.
Сравнение классификаций жизненных стратегий с делением видов на ценофилов и ценофобов
Основное различие этих классификаций — логическое основание деления. Классификации жизненных стратегий имеют логическим основанием особенности их биологии (аутоэкологические = систематические). Аналогичные классификации видов: классификация жизненных форм, морфологические и таксономические классификации. Деление же на ценофобы и ценофилы проводят по их положению в ЦС (т. е. логическое основание классификации — синэкологическое): ценофилы включены в ЦС, ценофобы — нет.
Виды, не входящие в состав ЦС, могут поселяться только в местах, где эти ЦС были разрушены в силу каких-либо причин. На практике это означает, что данные виды должны быть ярко выраженными r-стратегами. Виды поздних стадий сукцессионных рядов и климакса некоторых (преимущественно — наземных) ЦС обычно обладают качествами K-стратегов. Поэтому может создаться впечатление, что r-стратеги — это ценофобы, а K-стратеги — это ценофилы. Однако такое соответствие вовсе необязательно. Виды ранних сукцессионных стадий также являются r-стратегами. В сообществах, где происходит изъятие существенной части биомассы и особей доминирующих видов, даже виды климакса обладают свойствами r-стратегов. Примерами таких сообществ являются сообщества степей (Мордкович, 1982) и бентоса южного и среднего Каспия (Карпинский, 2001). В степях климаксные виды способны к быстрому восстановлению после стравливания. Бентос Каспия способен восстанавливаться после почти полного выедания рыбами. Для многих беспозвоночных и рыб, а также для деревьев характерно увеличение плодовитости с ростом размеров особи. Размер же особей увеличивается с возрастом, поэтому в данных группах долгоживущие виды (свойство К-стратегов) обладают высокой плодовитостью (свойство r-стратегов) и тем большей, чем больше продолжительность жизни, т. е. имеет место конфликт характеристик. Вообще, такие свойства r-стратегов, как высокая плодовитость и высокая способность к расселению, присущи большей части организмов макробентоса независимо от их конкурентоспособности и жизненной стратегии. Эти черты присущи практически всем видам, доминирующим в донных сообществах, в том числе таким видам-эдификаторам, которые по остальным показателям являются типичными К-стратегами, как многолетние макрофиты и герматипные кораллы.