Биогеография общая и частная: суши, моря и континентальных водоёмов - 2017 год
Австралийская область - Схемы биогеографического районирования суши - Биогеография суши
При оценке сходства Австралийской и Новозеландской биот с биотами Южной Америки и других районов Гондваны в первую очередь, обычно, обсуждают время их разделения. При этом север Гондваны (Африка, Индостан, Мадагаскар, север Южной Америки) всегда находились вблизи экватора. Юг Южной Америки, Австралия и Новая Зеландия находились в умеренной зоне (холодная, а тем более полярная зоны отсутствовали). После разделения приполярных районов только два фрагмента были достаточно велики для существования разнообразных СС: Австралия и Южная Америка. На остальных фрагментах Гондваны часть исходной биоты должна была вымереть просто из-за их малых размеров. Это одна из причин отсутствия в Новой Зеландии многих таксонов, имеющихся в Австралии и Южной Америке.
Последующая геологическая история фрагментов Гондваны также была различна. Одни (Мадагаскар, Новая Зеландия) остались в изоляции, на их территории сформировались высокоэндемичные, но в целом сравнительно малоразнообразные биоты. В Южной Америке поддерживался достаточно заметный термический градиент “экватор-полюс” обеспечивая разнообразие СС материка. Австралия же, имея вдвое меньшую протяженность с юга на север, двигалась из умеренного пояса в тропический. Это вызвало формирование новых СС, функционирующих в более тёплых условиях, и рост общего разнообразия биоты материка. Наиболее ясно последствия такого преобразования прослеживаются на растительности. При продвижении Австралии к северу, леса Nothofagus, простиравшиеся, судя по палеоботаническим данным, от Южной Америки через Антарктиду до Новой Гвинеи (Хамфриз, 1988), были заменены субтропической растительностью, где Eucalyptus стал почти единственной родом деревьев, разные виды которого освоили самые различные местообитания, от уровня моря до верхней границы леса в горах, дав колоссальную вспышку видообразования (Вальтер, 1982): по разным данным 450—700 видов. Как указывает Г. Вальтер (1968: 263) “виды Eucalyptus хорошо приспособлены к совместному произрастанию со злаками, в связи с этим между влажными и сухими эвкалиптовыми лесами и открытыми саваннами с эвкалиптами существуют лишь небольшие различия”.
Можно полагать, что и в других таксонах происходили сходные процессы. СС с климаксными ассоциациями Nothofagus (так же как и другая растительность умеренной зоны) сохранились лишь на юго-востоке континента и Тасмании, а также в горах Новой Гвинеи и Новой Каледонии, где климат, если и изменился, то в наименьшей степени. Там же сохранились и многие другие таксоны умеренной зоны Гондваны, многие сохранились также и на юге Южной Америки и в Новой Зеландии, что провоцирует некоторых авторов выделять Антарктическую область (см. выше).
Дальнейшее продвижение Австралии на север сопровождалось постепенным её сближением с Азией и ныне они располагаются ближе друг к другу, чем когда бы то ни было. Необходимо подчеркнуть, что пролив между Новой Гвиней и Австралией неоднократно исчезал. Высыхали и проливы между Зондскими островами и Азией. Но колебания уровня моря никогда не были столь велики, чтобы осушать глубоководное море Банда и проливы, соединяющие его с Тихим и Индийским океанами. Азия и Австралия никогда не были частью единой суши. Поэтому положение биогеографической границы на месте глубоководных проливов выглядит совершенно естественным (см. также Главу 2).
Большая часть границ, проводимых биогеографами в данном районе, расположены между линиями Уоллеса и Lydekker (на Рис. 3.40 эти линии более жирные). Линия Уоллеса совпадает с восточной границей азиатского шельфа. Линия Lydekker — с западной границей австралийского шельфа. Острова, расположенные между этими границами, никогда не соединялись с материками и были заселены биотами Азии и Австралии, преодолевавшими проливы. Так как источники биот архипелага (Австралия и Азия) расположены на его разных концах, в нём установилась клина, постепенный переход между биотами этих двух материков. Необходимо отметить, что до настоящего времени сведения о естественных биотах отдельных островов далеки от полноты. Так, изучение в 2012 (sic!) авиафауны Obi, одного из крупнейших островов этого района, показало, что из 109 видов авиафауны острова, 14 ранее на острове не отмечали, а 4 принадлежат к ещё неописанным видам (Mittermeier, Cottee-Jones, 2013). Наличие клины и плохая изученность региона позволяет проводить границу по-разному в зависимости от использованных критериев (своих у каждого автора) и изучаемой группы. Некоторые, отчаявшись, предлагают весь регион считать переходным (Cox, 2001).
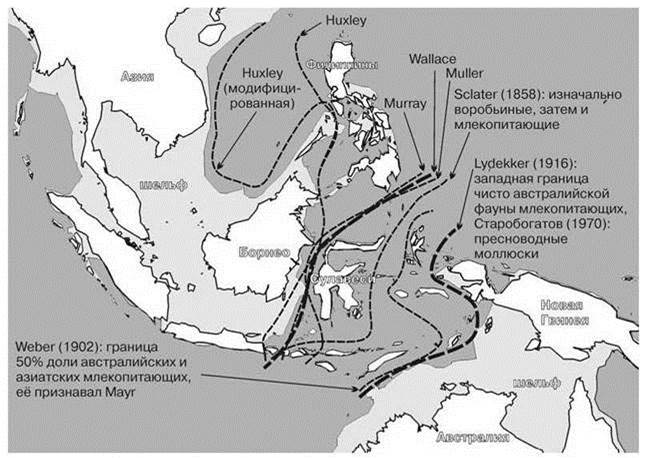
Рис. 3.40. Положение границы между австралийской и азиатской биотами по разным авторам, помещавших границу между Зондским архипелагом и Новой Гвинеей.
Авторы указаны на карте.
“Дождевые черви не считаются ни с линией Уоллеса, ни с моллукской линией [Вебера — ИЖ], ибо весь Моллукский архипелаг и даже Новая Гвинея заселены индийскими формами, австралийские же формы встречаются лишь за Торресовым проливом” (Пузанов, 1938: 334).
“Со времен Уоллеса (Wallace, 1876) было принято считать австралийскими многочисленные здесь [на Новой Гвинее — ИЖ] эндемичные таксоны, в том числе и те, которые отсутствуют в самой Австралии” (Крыжановский, 2002: http://www.zin.ru/Animalia/Coleoptera/rus/kry_pap.htm).
“По фауне большинства групп насекомых, например, стрекоз, богомолов, ряда семейств жуков (среди которых подробно изучены жужелицы, пластинчатоусые и листоеды), дневных бабочек, муравьёв, этот хорон хотя и занимает промежуточное положение между Австралийской и Индо-Малайской областями, всё же стоит заметно ближе к последней” (Крыжановский, 2002: 27—28).
“Большинство палеотропических мигрантов не смогло проникнуть на юго-запад Австралии, а большая часть общих [у Новой Гвинеи — ИЖ] с Австралией групп ограничена лишь юго-востоком Новой Гвинеи” (Крыжановский, 2002: 29).
“Различные австралийские животные проникают в районы открытых эвкалиптовых лесов южной части Новой Гвинеи, а новогвинейские животные проникают в районы дождевых лесов северо-восточной Австралии” (Дарлингтон, 1966: 378).
Рассмотрим как пример биоту острова Сулавеси, или Целебес, который Уоллес относил к Австралийской области. Остров площадью 174 600 км2 (11 по величине в мире) имеет горы высотой до 3478 м. Он отделён проливами с глубинами свыше 1000 м от остальных островов, имеет богатую фауну плацентарных млекопитающих (Geer et al., 2010). На острове в плейстоцене (лучше известна фауна раннего плейстоцена) жило много истреблённых видов азиатского происхождения, в том числе: два семейства слонов (настоящий слон Stegoloxodon celebensis и карликовый стегодон Stegodon sompoensis), гигантская свинья Celebochoerus heekereni, кабан Sus celebensis, два вида карликовых (<1 м в плечах) быков (Bubalus depressicornis и B. quarlesi), три вида макак и гигантская черепаха Testudo margae. Из уцелевших эндемов широко известна поныне свинья бабирусса с очень своеобразными клыками самцов (Рис. 3.41)**. С другой стороны эндемичный вид сорной курицы принадлежит к таксону, попавшему на остров с востока. В отличие от своих австрало-новогвинейских предков этот вид не строит инкубаторов из прелой листвы, а закапывает яйца в тёплый вулканический песок. Есть здесь и сумчатые млекопитающие. Поэтому остров относят то к Австралийскому царству, то к Палеотропическому.

Рис. 3.41 Череп бабируссы Babyrousa babyrussa. По: Wallace (1869).
Даже биота Филиппин носит отчётливый островной и отчасти переходный характер. Немногочисленные крупные виды млекопитающих (буйвол, кабан и три вида оленей) эндемичны. Свободные ресурсы используют и таксоны австралийского и новогвинейского происхождения: многие рода птиц общи с Австралийской областью, в частности здесь обитают сорные куры (заселившие даже Борнео), австралийские попугаи и растут эвкалипты.
Фактически линия Wallace отделяет острова Зондского архипелага, в периоды регрессий моря имевших сухопутную связь с материком с богатой азиатской биотой от обеднённых биот островов, никогда не соединявшихся ни с Азией, ни в Австралией. То же относится и к линии Lydekker. При этом в Новой Гвинее многие индо-малайские таксоны вновь представлены большим числом видов.
Общепринятое объяснение положения границы на месте Макассарского пролива между Борнео и Сулавеси, возникшего в эоцене, примерно позже 45 млн. лет назад (Lohman et al., 2011) и ныне имеющего ширину 200 км и глубиной до 2,5 км, восходящее к идеям А. Wallace, и которое считают несомненным авторы современных учебников биогеографии (см., например, Brown, Lomolino, 1998; Лебедева, Дроздов, Криволуцкий, 2004). Это объяснение удовлетворительно соответствует наблюдаемой картине только в случае некоторых таксонов наземных животных (млекопитающих и наземных моллюсков), некоторых птиц и пресноводную биоту (см. Главу 4).
В то же время анализ распространения растений (Вальтер, 1968; Разумовский, 1999), насекомых (Крыжановский, 2002) и рептилий выявляет гораздо большее сходство биоты Новой Гвинеи с Азией, нежели с Австралией. Эти авторы относят Папуасскую подобласть к Палеотропису. Даже новогвинейская фауна таких таксонов как земноводные, которым, казалось бы, сложно преодолевать морские проливы, более сходна с азиатской, нежели с Австралийской. Примером такой группы могут служить узкоротые квакши Microhylidae (Рис. 3.42), заселению ими Новой Гвинеи и Австралии способствовало то, что обитающие там виды не имеют стадии головастика, лягушки непосредственно развиваются из яиц. Другой пример — саговники рода Cycas (Рис. 3.43). Среди растений подобные ареалы (включающие Палеотропис или ещё более широко распространённые, Новую Гвинею и северо-восточную Австралию) широко распространены, ими обладают, например семейства: ивовые Salicacea, кокаиновые Erythroxylacea,непентесовые Nepenthacea (Рис. 3.44), Clusiaceae, Dichapetalaceae, Putranjivaceae, Hugonioideae, Malpighiaceae, Chrysobalanaceae. Много таксонов животных также обладают таким ареалом: нектарницы Nectariniidae (15 родов, 132 вида), питты Pittidae (1 род, 30 видов), райские птицы Paradisaeidae (Рис. 3.45). Много таксонов обладают сходными ареалами, но отсутствуют в Австралии, например характернейшие для тропических дождевых лесов диптерокарповые Dipterocarpaceae (Рис. 3.46), другие примеры семейств растений Ctenolophonaceae, Ixonanthaceae, Lophopyxidaceae, Pandaceae. Среди животных можно назвать птиц-носорогов Bucerotidae (14 родов, 57 видов) и хохлатых стрижей Hemiprocninae (1 род с 4 видами). Некоторые отличия Австралийской области и Новогвинейской подобласти Палеотрописа суммированы в Табл. 3.6.

Рис. 3.42. Ареал узкоротых квакш Macrohylidae
(413 видов, в 69 родах) (по http://en.wikipedia.org/wiki/Microhylidae).
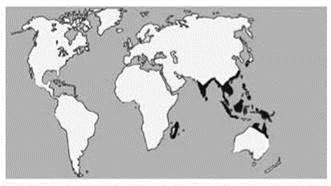
Рис. 3.43. Ареал саговников рода Cycas. По Nagalingum et al. (2011).
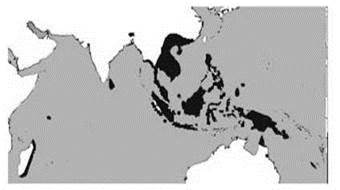
Рис. 3.44. Ареал непентовых Nepenthaceae. 1 род, 130 видов.

Рис. 3.45. Ареал райских птиц Paradisaeidae.
14 родов, 41 вид (в Австралии — 4 вида). По http://ibc.lynxeds.com/family/birds-paradise-paradis-aeidae.

Рис. 3.46. Ареал диптерокарповых Dipterocarpaceae в Старом свете. По Appanah, Turnbull (1998).
Табл. 3.6. Некоторые специфические черты Австралийской области и Новогвинейской подобласти Палеотрописа.
Особо отмечено для Новогвинейской подобласти нахождение таксона на северо-востоке Австралии (СВА).
|
климакс |
Австралийская область |
Новогвинейская подобласть |
Палеотропис без Новогвинейской подобласти |
|
субтропики, широко распространены мозаичные ЦС |
влажный тропический лес |
||
|
эвкалипты |
есть |
нет |
нет |
|
много разных семейств растений: ивовые Salicacea, кокаиновые Егуthroxylacea, непентесовые Nepenthaсеа, Clusiaceae, Dichapetalaceae, Putranjivaceae, Hugonioideae, Malpighiaceae, Chrysobalanaceae |
нет |
есть (СВА) |
есть |
|
много разных семейств растений: диптерокарповые Dipterocarpaceae, Ctenolophonaceae, Ixonanthaceae, |
нет |
есть |
есть |
|
Lophopyxidaceae, Pandaceae саговники Cycas |
нет |
есть (СВА) очень |
есть |
|
дневные бабочки |
малоразнообразны |
разнообразны (СВА) |
разнообразны |
|
птицы-носороги Bucerotidae |
нет |
есть |
есть |
|
хохлатые стрижи Hemiprocninae |
нет |
есть |
есть |
|
лягушки Microhylidae |
нет |
есть (СВА) |
есть |
|
настоящие лягушки Raninae |
нет |
есть (СВА) |
есть |
|
крокодилы |
нет |
есть (СВА) |
есть |
|
удавы Boinae |
нет |
есть (СВА) |
есть |
|
нектарницы Nectariniidae |
нет |
есть (СВА) |
есть |
|
питты Pittidae |
нет |
есть (СВА) |
есть |
|
райские птицы |
нет |
есть (СВА) |
нет |
|
страусы |
эму Dromaeidae 2 вида |
казуары Causariidae 6 видов (СВА) |
африканские страусы Struthionidae 1 вид |
|
древесные кенгуру |
нет |
есть (СВА) |
нет |
Поэтому здесь к Австралийской области отнесены только сама Австралия и то за исключением её севера, тогда как Новая Гвинея и север Австралии в качестве отдельной подобласти включены в Палеотропис. Границы Австралийской области и Новогвинейской подобласти Палеотрописа в нашем понимании совпадают с границами ботаника Р.В. Камелина (2012) и зоолога позвоночных Н.А. Бобринского (1946, 1951), последний, правда относил Новогвинейскую подобласть к Австралийской области.
Биогеография региона позволяет восстановить вероятное развитие событий. Первоначально Австралия располагалась в умеренном и субтропических поясах. Продвижение её к северу привело к тому, что северные районы Австралии оказались в тропическом поясе, т. е. к необходимости формирования тропических СС.
На территории Зондского архипелага и прилегающих районов Австралии можно выделить три типа СС: тропических лесов, мозаичные и горные (туманный, или моховой лес, полувечнозелёный лес) (Рис. 3.47). В Палеотрописе тропические леса занимают колоссальные площади, весьма разнообразны биотически и имеют большой возраст. В Новой Гвинее они не растут на юге и в горах, а в Австралии занимают сравнительно небольшие районы на северо-востоке. Когда в Палеотрописе формировались СС тропических лесов, Новой Гвинеи ещё не было, а Австралия располагалась в зоне субтропиков. Новая Гвинея начала возникать, сначала как небольшие острова, в раннем миоцене, примерно 20 млн. лет назад (Lohman et al., 2011). Напомню, что тропический дождевой лес сформировался в эоцене, более 10 млн. лет до этого. Поэтому, несмотря на определённые трудности с преодолением морских проливов, биота новогвинейских островов, была сформирована таксонами азиатских тропических лесов. Австралийские корни имеют только таксоны, аналоги которых не смогли преодолеть водные преграды. Во многих случаях корни таких таксонов, хотя и австралийские, но столь древние, что Новую Гвинею можно рассматривать как вполне самостоятельный центр возникновения и разнообразия. Примером такой группы могут служить райские птицы Paradisaeidae: 14 родов, 41 вид, в Австралии — 4 вида (Рис. 3.45) и казуары: 3 вида, все виды обитают на Новой Гвинее из них один также в Австралии. И райские птицы, и казуары в Австралии обитают в тропических лесах Индонезийской провинции (Рис. 3.50Б), относимых Разумовским (Рис. 3.46) к азиатской Малайской области.
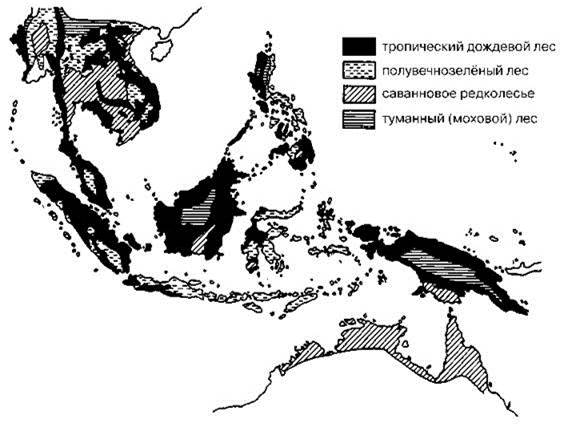
Рис. 3.47. Растительность юго-восточной Азии и северной Австралии.
Зато биота СС мозаичных районов, которые отсутствуют на Зондском архипелаге, но есть к югу от новогвинейских гор, в значительной степени австралийская по происхождению. Только здесь на Новой Гвинее встречаются столь характерные для Австралии эвкалипты, которые, смогли расселиться и на биотически наиболее обеднённые острова Палеотрописа, Филиппины в частности. Возникавшая периодически сухопутная связь с Австралией приводила к взаимному обмену: богатая азиатсконовогвинейская тропическая биота вселялась в районы, занятые тропическими лесами Австралии, распространение же субтропической биоты Австралии в значительной степени ограничено югом Новой Гвинеи. Результатом этой истории и является современная биогеография региона. Граница между ТДЛ с господством индо-малайских таксонов растений и районами с господством австралийских таксонов в Австралии “выражена очень резко как физиономически, так и флористически” (Вальтер, 1968: 261). Не удивительно, что ещё Альфред Уоллес, а вслед за ним и многие другие разделяли Австралийскую область на две подобласти: собственно Австралийскую подобласть и Папуасскую, к которой относят тропические районы Австралии даже зоологи позвоночных (Бобринский, 1946, 1951). Биогеографическое районирование, отражающее распределение СС разных зон было предложено С.М. Разумовским (Рис. 3.48). С Азией объединяет Новую Гвинею и Крыжановский (2002) по фауне насекомых и Белышев и Харитонов (1981) по фауне стрекоз. Последние, впрочем, больше связаны с континентальными водоёмами, нежели с наземными СС.
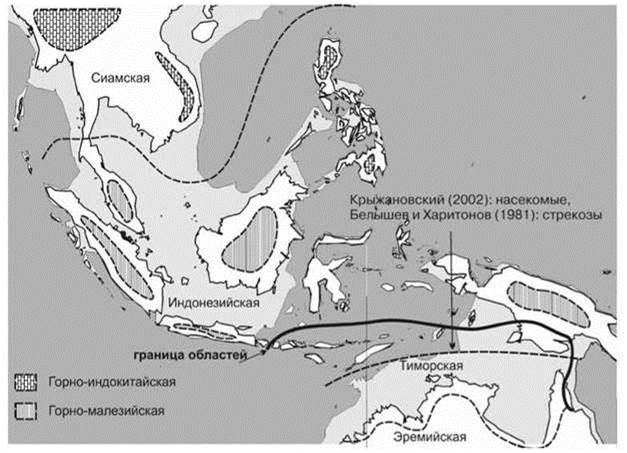
Рис. 3.48. Положение границы между австралийской и азиатской биотами по разным авторам, относившим Новую Гвинею к Палеотропису.
Районирование скомпилировано мной по оригинальным картам из архива С.М. Разумовского, подписаны названия провинций, их границы даны пунктиром; Тиморская и Эремийская провинции принадлежат австралийским областям, остальные — Малайской области; горные и эремийская — провинции субтропических лесов, остальные — тропических. Горные провинции показаны схематично.
Немалую роль в различии во взглядах на биогеографическое положение Новой Гвинеи играет и использование разных методов районирования. Если методом сравнения списков биот сравнивать списки географических регионов (остров Новой Гвинеи vs. материк Австралия), то по некоторым группам их сходство велико, что оправдывает объединение их в один регион. Если же сначала провести выделение биотически однородных районов, а затем сравнивать их биоты, то результаты получаются иными даже в отношении птиц и млекопитающих и это при том, что сухопутная связь Новой Гвинеи с Австралией прервалась только после повышения уровня Мирового океана всего 10 000 лет назад.
* * *
Австралия ассоциируется у людей с сумчатыми животными, недаром и на её гербе изображен кенгуру. Широко распространено мнение о том, что адаптивная радиация сумчатых стала возможной благодаря тому, что в Австралии никогда не было плацентарных животных. В действительности это не совсем так, вернее, совсем не так. Во-первых, и сейчас в состав австралийской биоты входит много видов плацентарных животных, попавших сюда до проникновения человека: это рукокрылые и грызуны. Число видов грызунов Австралии (67) всего в два раза меньше числа видов сумчатых (119). Все грызуны принадлежат к Muridae (настоящие крысы и мыши). Естественный ареал Muridaeограничен исключительно Старым Светом, но для ИндоМалайского архипелага характерна их сильная радиация, приводящая к образованию олиго- и монотипичных родов (Кузнецов, 2006). Хотя грызуны — сравнительно недавние вселенцы, они весьма длительное время сосуществуют с сумчатыми: об этом свидетельствует то, что грызуны и сходные с ними по размерам сумчатые потребляют разные ресурсы: среди мелких сумчатых нет грызущих видов, все они травоядные или насекомоядные. Связано ли это с вытеснением таких видов грызунами или ограниченными возможностями сумчатых к адаптации***, неясно. Во-вторых, по одному нижнему моляру описана Tingamarra, датированная границей палеоцена—эоцена (54,6 млн. лет назад). Этот зуб очень сходен с зубами южноамериканских копытных. Однако нахождение их в Австралии столь существенно меняет принятые ныне представления об эволюции австралийской биоты, что желательно получение дополнительных данных (Rose, 2006).
Среди остальных наземных позвоночных Австралии только черепахи содержат эндемов, ранга, аналогичному млекопитающим. Фауна земноводных и пресмыкающихся является результатом многократных инвазий из Азии с последующей адаптивной радиацией. Это подтверждает также отсутствие в этих таксонах особых связей с Южной Америкой или Африкой. Это и неудивительно, поскольку рептилии и земноводные в тропиках гораздо разнообразнее, чем в умеренных и субтропических широтах, где первоначально располагалась Австралия.
Что касается птиц, то ситуация с ними отлична от наземных позвоночных — они весьма разнообразны, а некоторые из возникших в Австралии таксонов в дальнейшем широко распространились по суше. Так, Corvini (вороны, галки, грачи и т. д.) возникли в Австралии, проникли в Азию в олигоцене—миоцене, откуда расселились по всем материкам. В Австралийской области расположен один из двух центров эволюции попугаев (другой — Неогея).
Как курьёз можно отметить живородящего сцинка Trachysaurus rugosus, обладающего настоящей аллантоидной плацентой (Пузанов, 1938).
* * *
Древнейшие останки человека в Австралии датируют 42 000 лет назад и собрали их в южной Австралии, поэтому, несомненно, человек достиг Австралии гораздо раньше. Древнейшие следы деятельности человека датируют 50 000 — 46 000 лет назад (Lewin, 2005), а, возможно и раньше. Человек застал Австралию значительно отличающуюся от современного её состояния.
Из-за малых размеров континента и отсутствия высоких гор достаточное для произрастания растительности количество осадков выпадает везде, поэтому девственная Австралия была покрыта сплошным растительным покровом, местами очень пышным, а не зарослями буша, злаковники и пустыни как ныне. Мегафауна была разнообразна, но большая часть к настоящему времени истреблена. Результатом этого была кардинальная перестройка СС. До появления человека значительные территории были заняты лесами хвойных (араукариями), ныне полностью уничтоженными. В умеренных районах юга континента и Тасмании климаксные ассоциации — леса Nothofagus.
Этот процесс детально изучен по динамике спор в кратере Lynch на северо-востоке Австралии (Rule et al., 2012). К моменту появления здесь людей (примерно 49 000 лет назад) растительность была образована примерно поровну дождевым лесом, образованным голо- и покрытосеменными и склерофильной растительностью. Злаки почти отсутствовали. Мегафауна была многочисленна (судя по наличию спор Sporormiella, гриба специфичного для помёта крупных животных). Примерно 41 000 лет назад сначала почти исчезли споры Sporormiella и возросло количество частиц угля (ранее практически отсутствовавших), затем возросло обилие пыльцы злаков и склерофильной растительности с одновременным уменьшением пыльцы деревьев дождевого леса и в первую очередь пыльцы голосеменных. Все изменения заняли <1000 лет. Авторы объясняют эти изменения деятельностью человека, истребившего мегафауну и выжегшего дождевой лес. Результатом этой перестройки было широкое распространение злаковников и склерофильной растительности.
“Местные жители ещё до появления белых поселенцев так часто пускали палы..., что это способствовало изменению облика некогда облесённых территорий” (Вальтер, 1968: 203).
“Равнина Налларбор в настоящее время лишена почвенного покрова, последний... был развеян и перемещён в восточном направлении” (Вальтер, 1968: 444).
“Во внутренних областях Австралии отсутствуют пустыни, существование которых обусловлено климатом. Если тем не менее на географических картах Австралии к северу от озера Эйр расположена область, которую называют “пустыня Симпсон”, а западнее от неё — ещё большая по площади “пустыня Гибсона”, то это объясняется расхождениями в определении понятия пустыня” (Вальтер, 1968: 437).
Австралия расположена южнее пассатов, в более сухом климате, в котором пожары распространяются гораздо легче. Особенно выжигание преобразовало, естественно, менее увлажнённые территории, вызвав широкое распространение ксеро- и литосерий, местами приведя к полному уничтожению растительности, а затем и почвы в результате эрозии, преимущественно ветровой. В умеренных районах с хорошим увлажнением (Тасмания и юго-восток Австралии) выжигание привело к замене климаксных лесов Nothofagus пирогенными стадиями с доминированием эвкалиптов или верховых болот (Вальтер, 1974). О том, что климаксными являются именно леса Nothofagus, а не леса эвкалиптов, которые растут там сейчас, свидетельствует то, что подрост эвкалиптов отсутствует под пологом Nothofagus, тогда как Nothofagus хорошо возобновляются в эвкалиптовых лесах (что характерно для демутационной сукцессии). Система человек → пожары → уничтожение мегафауны → опустынивание действовала, таким образом, как обычно, с положительной обратной связью и привела к формированию малопродуктивного буша, занимающего большую часть Австралии и так называемых “пустынь”. И это при том, что в Австралии, в отличие от других биогеографических областей Земли, отсутствуют условия для возникновения настоящих пустынь. Хотя вдоль её западных берегов проходит холодное течение, здесь нет постоянного обширного апвеллинга, создающего береговые пустыни Намибии и Перу. Косвенным подтверждением отсутствия в области естественных пустынь и очень молодой (вероятно, антропогенный) возраст современных территорий, со скудной растительностью или вовсе её лишённых, служит полное отсутствие в Австралии суккулентов (Вальтер, 1968: 300). Богатство австралийской растительности, как и ТДЛ поддерживалось замкнутым круговоротом. Неизбежным следствием нарушения таких СС (вырубка, пожары) была эрозия, уносящей почву и биогены. Потеря биогенов привела к резкому падению продуктивности. Для восстановления их прежнего количества требуются как минимум тысячелетия. Положение резко ухудшается на старых платформах и кристаллических щитах, где нет вулканов и которые сотни миллионов лет не покрывались морем. В этом случае единственные источники биогенов — это принос животными и медленная эрозия кристаллических пород (гранитов и т. п.). Поэтому утраченные в результате эрозии почв биогены фактически не способны появиться вновь. Это означает, что если произошла замена австралийских лесов “пустынями”, то без вмешательства человека прохождение сукцессий и достижение климакса невозможно (биотопический субклимакс). Это подтверждает низкая скорость восстановления лесов, и другой естественной растительности после того, как она была уничтожена. Рентабельность сельского хозяйства Австралии очень мала по той же причине: посевы зерновых требуют много удобрений, а приемлемая нагрузка на пастбища очень низка (Diamond, 2005).
Из-за сильного антропогенного воздействия на протяжении последних 50 000 лет распределение современной растительности Австралии (Рис. 3.49) сильно отличается от естественного.
Сравнение карты геоботаника Г. Вальтера (Рис. 3.49) со схемами биогеографов Brown, Lomolino (1998) (Рис. 3.50 А) и схемой распределения СС, составленной С.А. Курганской (Рис. 3.50 Б) (Разумовский, Лабунцова, 1969) наглядно иллюстрирует, как подход к составлению карты предопределяет результат.
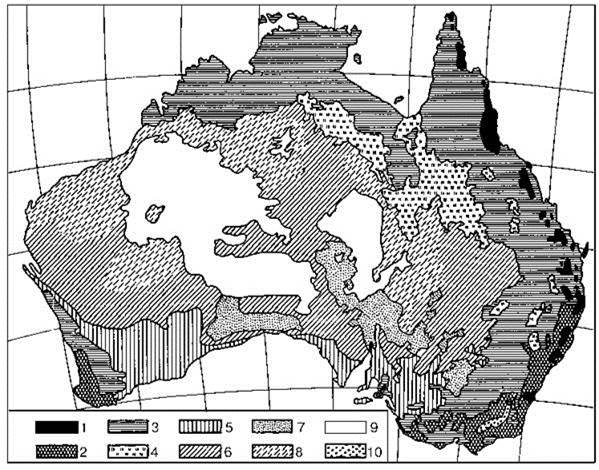
Рис. 3.49. Карта растительности Австралии по Г. Вальтеру (1986).
1 — тропически-субтропический дождевой лес с индо-малайскими флористическими элементами; 2 — влажный эвкалиптовый лес; 3 — сухой эвкалиптовый лес; 4 — злаковники, образованные Astrebla; 5 — разреженные заросли кустарниковых эвкалиптов (мали-скраб); 6 — заросли Acacia aneura и др. (мульга); 7 — солянковая полупустыня, образованная видами Atriplex и Kochia; 8 — сообщества злаков с колючими листьями (спинифекс Triodia); 9 — необитаемые области называемые “пустынями”; 10 — альпийские луга (только на юго-востоке, в Австралийских Альпах).

Рис. 3.50. Биогеографическое районирование Австралии.
А — биогеографические провинции по Brown, Lomolino (1998): 1 — Торреская; 2 —Басская; 3 — озера Эри; 4 — Вестралия.
Б — ботанико-географические провинции по С.А. Курганской из Разумовского, Лабунцовой (1969). Провинции: тропических лесов: 1 — Индонезийская, 2 — Тиморская; субтропических лесов: 3 — Эремейская, 4 — Западноавстралийская, 5 — Восточноавстралийская, 6 — Тасманийская.
Схема Г. Вальтера отражает распределение различных типов растительности в данный момент без учёта видового состава, сукцессионных тенденций и антропогенных изменений. Схемы Brown, Lomolino и С.А. Курганской, напротив, отражают типы экосистем и биотический состав. Brown, Lomolino пишут: Торресская провинция населена “животными и растениями, близкими к Новогвинейским и, иногда, ЮгоВосточной Азии” (стр. 303), провинция озера Эри — это аридные и полуаридные экосистемы, юго-восток Австралии населена животными и растениями умеренного климата с Nothofagus, для Вестералии характерен высокий эндемизм и некоторое сходство с южной Африкой, в частности наличием семейства растений Proteacea”. Схема С.А. Курганской отличается большей детальностью (Торесская и Басская провинции Brown и Lomolino разделены каждая на две) и гораздо большей извилистостью границ. И то, и другое стало возможным благодаря использованному методу выделения конкретных биот, тогда как схема Brown и Lomolino является, по-видимому, их экспертной оценкой и границы проведены на глаз.
Ниже перечислены наиболее известные представители биоты девственной Австралии, ныне исчезнувшие (Рис. 3.51).
Diprotodontidae. 6 ископаемых родов. Diprotodon несколько видов, D. australis достигал 3 м длины и 1,8 м высоты. D. optatum — 3,8 м длины и 1,7 м высоты. Вымерли 50—18 тысяч лет назад. Самый крупный Zygomaturus — Z. trilobus — размером с буйвола или карликового бегемота, весом 300—500 кг, вероятно околоводный, вымер примерно 50000 лет назад. Последний представитель рода вымер 15000 лет назад.
Palorchestes azeal представитель отдельного семейства Palorchestidae. Как тапиры и южноамериканские лошади он имел короткий хобот. Достигали размера лошади.
Кенгуру. Всего в плейстоцене вымерло 6 родов кенгуру (Дарлингтон, 1966). Коротколицые кенгуру Procoptodon достигали роста ~3 м. На скакательных задних ногах было по одному пальцу. Наиболее известен P. goliah. Относящийся к тому же семейству, что живущие ныне крупные виды кенгуру род Propleopus включал два плейстоценовых вида: P. oscillans и P. wellingtonensis. В отличие от остальных кенгуру, они, вероятно, были хищными.
Genyornis newtoni — последний представитель эндемичных Dromornithidae, страусоподобных нелетающих птиц. Рост его — 2 м, вес — 200—240 кг. Обитал в лесах и степях Австралии 50—30 тысяч лет назад. Исследование изотопов яичной скорлупы G. newtoni и страуса эму показало, что 130—59 тысяч лет назад эму ели траву, кустарник и деревья, а G. newtoni исключительно траву. Примерно 50 тысяч лет назад эму перестали есть траву, а G. newtoni вымерли.
Мейолании — огромные сухопутные черепахи, экологические аналоги Testudinidae. Этих черепах относят к иному подотряду, нежели всех остальных современных черепах (сухопутных, пресноводных и морских). Этот подотряд возник, вероятно, в триасе и был доминирующим в мезозое, но позднее эоцена известен только из Австралии и прилежащих островов (2 рода в самой Австралии плюс несколько более мелких островных видов). Самая крупная Meiolania имела карапакс длиной около 2,0 м (Rhodin et al., 2015), её вес, вероятно, превышал 1 т. На голове мейоланий были мощные рога, они имели длинный хвост (редкое явление у черепах) вооружённый бронированными кольцами и шипами. Дольше всего, до XVII—XIX вв. просуществовали мейолании на островах Лорд-Хау (Lord Howe, остров площадью в 13 кв. км в 550 км к востоку от Австралии) и Уолпол (Walpole, к юго-востоку от Новой Каледонии, площадью 2 км2) и на Новой Каледонии, но вскоре после открытия их человеком и там от них остались лишь кости и воспоминания.
“Australia’s megafauna included 20 or more genera of giant marsupials, monotremes, birds, and reptiles (Johnson, 2006), which were extinct by 40 thousand years ago (Roberts et al., 2001; Miller et al., 2005; Gillespie et al., 2006; Grun et al., 2008), soon after people colonized Australia, suggesting that people caused the extinction. Proposed mechanisms are overhunting (Johnson, 2006; Brook, Johnson, 2006), vegetation change due to landscape burning by people (Miller et al., 2005), or a combination thereof” (Rule et al., 2012: 1484).
В состав крупнейших наземных хищников входили, естественно, сумчатые. Самый крупный сумчатый хищник — Thylacoleo carnifex, достигал размеров современного ягуара: длина от кончика морды до основания хвоста — 110 см, высота в холке — 70 см, вес — около 100 кг. Детали скелета сумчатого льва дают основания предположить, что животное могло садиться на задние лапы, опираясь на сильный хвост, как это делают кенгуру, и залезать на деревья на манер леопарда. Интересен противопоставленный большой палец. Вымер 40—30 тысяч лет назад, после появления в Австралии человека, обнаружены наскальные рисунки Th. carnifex (Akerman, Willing, 2009), на которых он изображён полосатым подобно тигру. Возможно, ещё более крупным хищником были уже упомянутые кенгуру Propleopus. Но самыми крупными хищниками Австралии были рептилии.
Megalania prisca — огромный австралийский варан (многие и включают его в род Varanus) длиной 5—7 м (по другим данным до 9 м) и 700—1500 кг весом. Вымер около 40 тысяч лет назад.
Хотя все современные крокодилы не могут долго обходиться без водоемов, где проводят почти всю свою жизнь, на рубеже кайнозоя и мезозоя, когда крупные хищные динозавры вымерли, а хищные млекопитающие ещё не возникли, их роль в Африке, Европе и Южной Америке выполняли сухопутные крокодилы, но все они вымерли миллионы лет назад. Лишь в Австралии и прилежащих островах они не просто сохранились, но были разнообразны. Здесь в плейстоцене жило по меньшей мере четыре рода сухопутных бегающих крокодилов — Quinkana, Pallimnarchus, Volia и Mekosuchus. Первые два рода найдены в самой Австралии; последние обитали на островах Южной Пацифики, таких как Фиджи, Вануату и Новая Каледония. Сухопутные крокодилы вели образ жизни, сходный с образом жизни современных варанов, соответственно и зубная система у них была иная, нежели у современных водных крокодилов, это позволяло им откусывать куски добычи, а не выкручивать. Некоторые виды, возможно, предпочитали питаться прибрежными моллюсками и были сравнительно мелкими — 2—3 м в длину, хотя в плиоцене встречались и семиметровые. В плейстоцене вымирание сухопутных крокодилов происходило после появления в данной местности человека. В Австралии они вымерли примерно 40 тысяч лет назад (возможно немногим позже); на Новой Каледонии (там обитал только Mekosuchus inexpectatus) — примерно в 1670 г. н. э. (Naish, 2009).
Wonambi — наземные змеи, достигавшие 6 м в длину, 30 см в поперечнике и до 600 кг весом. Принадлежали к вымершему семейству Madtsoiidae. Известно два вида. Их биология была, вероятно, сходна с удавами. Вымерли в последние 50000 лет.
Расположенный рядом с Австралией небольшой (18 575 км2) остров Новая Каледония имеет высокоэндемичную флору: пять эндемичных семейств 108 эндемичных родов, из 3061 видов 80% — эндемы (Morat, 1993). Среди них некоторые весьма примитивны: половина (шесть из 12) живущих в настоящее время на Земле родов покрытосеменных растений без сосудов в стебле (не считая групп, где они вторично редуцировались) обитает здесь. Тахтаджян (1978, 1986) выделил остров в отдельное подцарство (хотя по его же собственным критериям это не более, чем провинция). Фауна не столь уникальна, хотя, как любой длительно изолированный остров, имеет ряд эндемичных видов. Современный эндемизм биоты, как обычно, уменьшился после заселения Новой Каледонии человеком, истребившим местную мегафауну. Из мегафауны можно назвать уже упоминавшихся крупных черепах, сухопутных крокодилов, а также гигантскую большеногую курицу Sylviornis neocaledoniae, общей длиной 1,7 м и весом 30 кг, инкубаторы, построенные этим видом, даже сейчас, спустя 3500 лет после истребления куриц, достигают 5 м высоты и 50 м в диаметре. Горные леса Новой Каледонии с Nothofagusзаслуживают выделения в отдельный биогеографический регион.

Рис. 3.51. Некоторые представители мегафауны, исчезнувшие с появлением человека в Австралии.
Рядом с Megalania для сравнения изображён варан о. Комодо. Рисунки с сайтов http://donsmaps.com, http://uk.wikipedia.org, http://australianmuseum.net.au/Australias-extinct-animals#.