Биология в экзаменационных вопросах и ответах для абитуриентов, репетиторов, учителей
Каковы основные формы естественного отбора с позиций синтетической теории эволюции - Эволюция живых систем - ОБЩАЯ БИОЛОГИЯ
В любом биогеоценозе противоречия между особями одного или разных видов разрешаются в борьбе за существование. Закономерным результатом борьбы за существование является гибель одних особей популяции и выживание и размножение других, т.е. естественный отбор. Он определяет, какие аллели будут переданы следующему поколению благодаря дифференциальным (избирательным) преимуществам, которые они обеспечивают, проявляясь в фенотипах.
В настоящее время выделяют три формы естественного отбора, — стабилизирующий, движущий, или направленный, и дизруптивный.
Стабилизирующий отбор — это форма естественного отбора, направленного на поддержание в популяции среднего, ранее сложившегося признака. Учение о стабилизирующем отборе разработал советский ученый И. И. Шмальгаузен.
Стабилизирующий отбор происходит в тех случаях, когда фенотипические признаки оптимально соответствуют условиям среды и конкуренция относительно слабая. Такой отбор действует во всех популяциях, уничтожая особей с крайними отклонениями признаков. В любой популяции в силу ее генетической разнокачественности появляются на свет особи с разной степенью выраженности того или иного признака. Такое разнообразие особей по любому признаку обеспечивается генетико-экологическими факторами, воздействующими на популяции в течение многих поколений. Если подсчитать количество особей, имеющих ту или иную выраженность данного признака, то окажется, что большинство будет приближаться к некой средней величине, средней норме. Менее всего будет особей, имеющих наибольшее отклонение от средней величины признака. Стабилизирующий отбор приводит к уничтожению крайних отклонений и как бы стабилизирует среднюю норму выраженности признака (рис. 37). Он наблюдается в том случае, если условия внешней среды длительное время остаются постоянными. В относительно неизменной среде преимуществом обладают типичные, хорошо приспособленные к ней особи со средним выражением признака, а отличающиеся от них погибают. Например, в 1898 г. американский орнитолог Г. Бампас после сильных ветров и снегопада обнаружил 136 оглушенных и полуживых домовых воробьев. При отогревании 72 из них выжили, 64 погибли. Оказалось, что погибшие воробьи имели или очень длинные, или очень короткие крылья; наиболее приспособленными в такой ситуации оказались воробьи со средней нормой реакции признака.
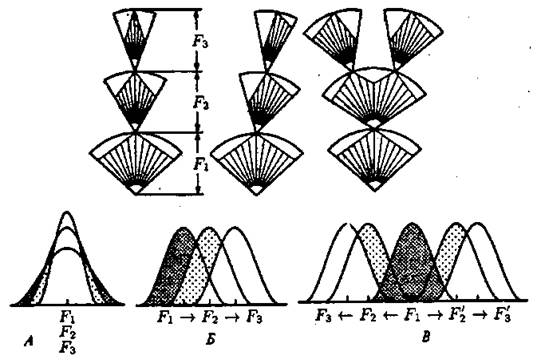
Рисунок 37. Схема действия стабилизирующей (А), движущей (Б) и дизруптивной (В) форм естественного отбора (по Н. В. Тимофееву-Ресовскому и др., 1977). F — поколения. На популяционных кривых заштрихованы элиминируемые варианты. Величина дуги при отборе внутри одного потомства соответствует норме реакции
Движущий отбор был описан еще Ч. Дарвином, а современное учение о движущем отборе разработано Дж. Симпсоном. Суть этой формы отбора заключается в том, что при медленном изменении условий среды в новом направлении неуклонно происходит сдвиг средней нормы в ту или иную сторону. Движущий отбор, таким образом, приводит к эволюционному изменению, оказывая на популяцию такое давление, которое благоприятствует увеличению в ней частоты новых аллелей (рис. 37). После того как новая средняя норма выраженности признака (средний фенотип) придет в оптимальное соответствие с новыми условиями среды, вступает в действие стабилизирующий отбор.
Классическим примером эволюционного изменения по типу движущего отбора является появление темноокрашенных форм некоторых видов бабочек под воздействием химических загрязнений атмосферы. За последние 100 лет у более 80 видов бабочек появились темные формы. Это явление известно теперь под названием индустриального (промышленного) меланизма. До 1848 г. все описанные формы бабочек березовой пяденицы имели бледно-кремовую окраску с черными точками и отдельными темными пятнышками. В 1848 г. в Манчестере была обнаружена черная форма этой бабочки, а к 1895 г. популяция березовой пяденицы в Манчестере состояла на 98 % из темноокрашенных бабочек. Эта меланическая форма появилась в результате возникновения случайных мутаций, причем фенотип мутантных особей обладал в промышленных районах большим преимуществом по сравнению со светлоокрашенными формами. Светлоокрашенные формы были незаметными на стволах берез, покрытых лишайниками. С интенсивным развитием промышленности диоксид серы, образующийся при сжигании угля, вызвал гибель лишайников в промышленных районах, и в результате обнажилась темная кора деревьев, которая сделалась еще более темной из-за покрывающей ее сажи. На темном фоне светлоокрашенные пяденицы склевывались малиновками и дроздами, а выживали и успешно размножались меланические формы, которые на темном фоне менее заметны.
Для многих видов характерен полиморфизм — существование двух или нескольких форм по тому или иному признаку. Полиморфизм нельзя объяснить только возникновением новых мутаций. Причины его могут быть разные. Он может быть обусловлен повышенной относительной жизнеспособностью гетерозигот. В других случаях полиморфизм может быть результатом действия особой формы движущего отбора, получившей название дизруптивною (разрывающего) отбора. Дизруптивный отбор — процесс расчленения ранее целостной популяции на отдельные формы (две и более адаптивные нормы) путем сохранения крайних вариантов фенотипов и элиминации промежуточных. Эта форма отбора осуществляется в тех случаях, когда две или более генетически различные формы обладают преимуществом в разных условиях, например, в разные сезоны года. Дизруптивный отбор благоприятствует более чем одному фенотипу и направлен против промежуточных форм. Он как бы разрывает популяцию по данному признаку на несколько групп, встречающихся на одной территории, и может при участии изоляции привести к разделению популяции на две и более (см. рис. 37). В основе дизруптивного отбора лежит дарвиновская дивергенция.
Моделью дизруптивного отбора может быть ситуация возникновения карликовых рас хищных рыб в малокормном водоеме. Часто щурятам-сеголеткам не хватает корма в виде мальков рыб. В этом случае преимущество получают самые быстрорастущие, которые очень быстро достигают размеров, позволяющих поедать своих собратьев. С другой стороны, в выгодном положении окажутся щурята с максимальной задержкой скорости роста, так как мелкие размеры позволяют им длительное время оставаться планктонофагами. Подобная ситуация через стабилизирующий отбор может привести к возникновению двух рас рыб.