Биология - Краткий курс с рисунками и коспект-схемами для студентов - А. Б. Чердак - 2016 год
ВЗАИМОДЕЙСТВИЕ МЕЖДУ ГЕНАМИ - Законы наследственности - Основы генетики
Генотип человека — тысячи различных признаков — размещается всего в 46 хромосомах. Это означает, что каждая хромосома содержит множество генов. Законы Менделя справедливы только для генов, локализованных в разных хромосомах. Гены, находящиеся в одной хромосоме, называются сцепленными. Все гены, относящиеся к одной группе сцепления, входят в одну хромосому и при образовании гамет наследуются вместе. Количество генов в различных группах сцепления (то есть в различных хромосомах) может отличаться друг от друга. При дигибридном скрещивании сцепленные гены, как правило, не подчиняются законам Менделя. С другой стороны, полное сцепление случается достаточно редко, и в потомстве обычно бывают представлены все четыре фенотипа. Таким образом, и в этом случае при дигибридном скрещивании образуются новые сочетания признаков — рекомбинантные фенотипы. Итак, если особи с новыми генными комбинациями встречаются в потомстве реже, чем особи с родительскими фенотипами, то это верный признак сцепленности соответствующих генов. Появление рекомбинантных сочетаний у аллелей называется кроссинговером (перекрестом).
Исследования генетиков начала XX века показали, что кроссинговер имеет место в результате разрыва и рекомбинации гомологичных хромосом и происходит практически между всеми хромосомами. Частота рекомбинаций определяется по формуле т = N/No, где N — количество рекомбинантов, NO — общее количество потомков. В то же время частота рекомбинаций определяет число рекомбинаций, происходящих при образовании гамет.
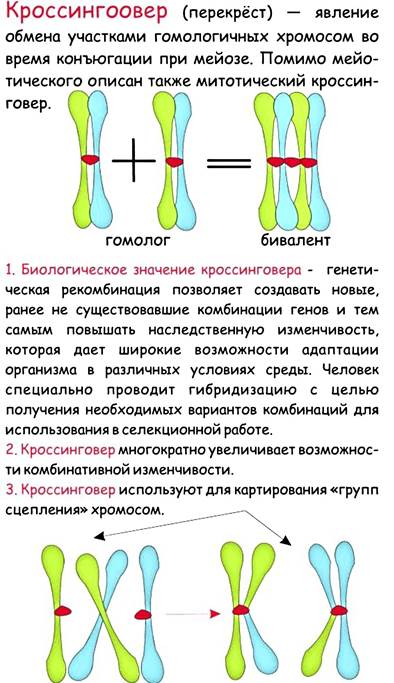
Частота рекомбинаций генов показывает относительное расположение сцепленных генов в хромосоме: чем дальше друг от друга находятся гены, тем выше частота рекомбинации. Это обстоятельство используется при составлении генетических карт. Условное “расстояние” между локусами (местоположениями в хромосоме) двух генов считается прямо пропорциональным частоте рекомбинации. Взаимное расположение (последовательность) локусов трёх и более генов определяется методом триангуляции. При этом сначала берутся гены с наименьшей частотой рекомбинации. Далее выбирают следующую по величине частоту рекомбинации и указывают два возможных положения нового гена; одно из этих положений будет отсеяно на следующем шаге, когда берётся третья частота.
В реальных экспериментах генетические карты могут искажаться благодаря двойному кроссинговеру, когда рекомбинация происходит одновременно в двух точках. Двойной кроссинговер особенно характерен для генов, локусы которых разделены большими расстояниями.
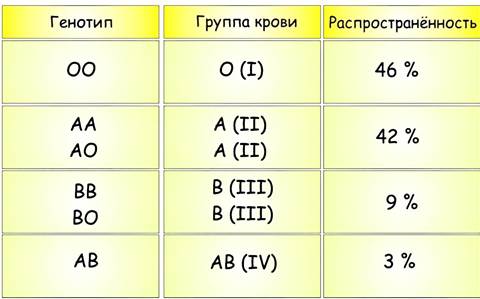
Существуют и более сложные механизмы взаимодействия между генами. Так, каждый признак может контролироваться не двумя, а тремя и более аллелями. Примером подобных множественных аллелей является наследование групп крови у человека. Три аллеля гена группы крови обозначаются буквами А, В и О. Аллели А и В являются доминантными, а аллель О рецессивен им обоим. В результате у человека могут наблюдаться четыре различные группы крови.
Не во всех генах можно указать доминантный и рецессивный аллели. Иногда в гетерозиготном состоянии ни один из аллелей не доминирует над другим. В этом случае говорят о неполном доминировании. Отношение фенотипов во втором поколении в этом случае отличается от менделевского 3:1; как правило, у половины особей сохраняется родительский фенотип.
Нетипичное менделевское соотношение может быть следствием не только неполного доминирования, но и наличия в генотипе организмов рецессивного гена. Если этот ген летальный (то есть вызывающий гибель его носителей), то организмы, гомозиготные по летальному гену, могут умереть ещё до рождения.
Ещё одним способом взаимодействия между аллелями является сверхдоминирование, когда определяемый геномом благоприятный признак проявляется у гетерозиготных организмов. Ряд учёных считает, что именно сверхдоминирование объясняет явление гибридной силы (гетерозиса), известное селекционерам еще с XVIII века: потомки скрещиваний между разными сортами растений или породами животных заметно превосходят родительские формы по большому числу признаков.
Один ген может влиять сразу на несколько признаков (явление плейотропного действия генов); так, мутация одного из генов дрозофилы приводит к рубиновой окраске глаз, уменьшению размеров тела, пониженной жизнеспособности и бесплодию. Наряду с этим, многие признаки определяются не одним геном, а целым генным комплексом — взаимодействием нескольких генов, находящихся, возможно, в разных локусах (далеко друг от друга). Ряд заметных признаков организма являются следствием воздействия многих генных комплексов (полигенной системы). Вклад каждого из генов в фенотип при этом достаточно мал, и можно говорить о непрерывной изменчивости организмов по данному признаку. В частности, один из генов (эпистатический ген) может подавлять эффект другого гена; в другом случае, один из генов может влиять на проявление другого гена (в этом случае говорят о комплементарности генов).
Интересным примером взаимодействия между генами являются признаки, связанные с полом. Как известно, гомологичные хромосомы во всех парах хромосом за исключением одной (гетеросом) идентичны друг другу. В гетеросомах заключены гены, определяющие пол особи. У самок имеются два идентичных гена (XX), у самца в гетеросомах гены разные (ХY). У некоторых групп животных наблюдается обратное соотношение (одинаковы гены у самца); наконец, у отдельных групп животных Y-хромосомы нет вообще.
Если у человека У -хромосома приводит к развитию семенников и становлению мужской особи, то у большинства животных Y-хромосома содержит очень мало генов и не имеет отношения к полу; мужские половые признаки у таких животных проявляются в присутствии одной Х-хромосомы, но маскируются при наличии пары Х-хромосом. Это пример наследования, ограниченного полом. Бывает и такая ситуация, что признак определяется геном, находящемся в Х-хромосоме, а в Y-хромосоме соответствующего ему участка нет вообще. Поэтому у особей мужского пола подобные признаки проявляются, даже если соответствующие гены рецессивны.