Биология - Учебно-практический справочник
Закономерности наследственности - Организменный уровень организации жизни
Методы работы Г. Менделя
Мендель проводил опыты на горохе посевном, который оказался удобным модельным организмом. Существует много сортов этого растения, отличающихся окраской семян, цветков, длиной стебля, структурой поверхности семян и т. д. Это растение, которому свойственно самоопыление, следовательно, потомки каждой особи являются генотипически однородными, гомозиготными по большинству генов - являются чистыми линиями. Горох можно искусственно опылять перекрестно, а это позволяет гибридизировать чистые линии. Мендель четко определял условия эксперимента — выделял среди многих наследственных признаков разные состояния одного или нескольких признаков и проследил их проявления в ряду поколений. В результате статистической обработки были объяснены закономерности передачи различных состояний наследственных признаков в ряду поколений гибридов.
Законы Менделя
При скрещивании двух гомозиготных организмов, отличающихся друг от друга по одной паре альтернативных (взаимоисключающих) признаков, все потомство в первом поколении единообразно и по фенотипу, и по генотипу (I закон Менделя).
В опытах Менделя при скрещивании сортов гороха, которые имеют желтые и зеленые семена, все потомство оказалось с желтым семенами. При этом не имело значения, какую окраску имели родительские растения. Стало ясно, что они в равной степени способны передавать свои признаки потомству. Аналогичные результаты были получены в других опытах. Данная закономерность получила название правила единообразия гибридов первого поколения. Признак, который проявляется в первом поколении, называется доминантным, а не проявляющийся — рецессивным.
Для объяснения этих результатов следует обратиться к данным хромосомной теории наследственности. Каждая клетка тела имеет диплоидный набор хромосом, в каждой из которых находятся разные аллели одного гена. То же самое происходит и в зиготе, где обычно находятся два аллеля одного гена и генотип, которые по какому-либо признаку можно записать двумя буквами. Гомозиготная особь по доминантной или рецессивной аллели обозначается соответственно АА или аа, а гетерозиготная — Аа. При полном доминировании рецессивный аллель проявляется только в гомозиготном состоянии, а доминантный — как в гомозиготном, так и в гетерозиготном состояниях.
При скрещивании двух гетерозиготных особей, которые анализируются по одной альтернативной паре признаков, среди потомков ожидается расщепление по генотипу 1 : 2 : 1, а по фенотипу (при полном доминировании) — 3 : 1 (II закон Менделя).
При скрещивании единообразных гибридов первого поколения между собой во втором поколении появляются особи и с доминантными, и с рецессивными признаками, т. е. возникает расщепление в определенных соотношениях: 75% особей имеют доминантные признаки, а 25% — рецессивные. Аллельные гены, находясь в гетерозиготном состоянии, не изменяют друг друга, при созревании гамет у гибридов образуется примерно равное количество гамет с доминантными и рецессивными аллелями; при оплодотворении мужские и женские гаметы свободно комбинируются. При скрещивании гетерозигот (Аа), следует ожидать четыре возможных сочетания: АА, Аа, Аа, аа. По фенотипу особи АА и Аа не отличаются, и проявляется расщепление 3 : 1. Однако по генотипу соотношение остается 1AA : 2Аа : 1aa.

При полигибридном скрещивании расщепление состояний каждого признака происходит независимо от других. Например для дигибридного скрещивания характерно расщепление 9 : 3 : 3 : 1 (III закон Менделя).
При полигибридном скрещивании родительские особи анализируются по нескольким альтернативным признакам. Простейшим примером является дигибридное скрещивание, проведенное между растениями двух сортов гороха с различной окраской и формой семени. Родители являются дигомозиготами по этим признакам (ААВВ и ааb). Первое поколение гибридов в этом случае оказывается единообразным, проявляются только доминантные признаки. Их генотип АаВb.

При скрещивании особей F1, гетерозиготных по двум признакам, расщепление — 9 : 3 : 3 : 1. Чтобы объяснить полученный результат, следует учитывать, что при мейозе у дигетерозиготных особей первого поколения образуется 4 типа гамет: АВ, Bb, аВ, ab. При скрещивании этих особей возможны 16 комбинаций генотипов. Их легче всего определить, используя решетки Пеннета.
Анализируя наследование при дигибридном скрещивании каждого признака отдельно, можно убедиться, что оно соответствует первому и второму законам Менделя. Это означает, что при полигибридном скрещивании различные признаки комбинируются независимо. Расщепление при этом получается (3 : 1), где n — количество анализируемых признаков.
Как дополнение к законам Менделя часто используют закон чистоты гамет, сформулированный в 1902 г. В. Бетсоном. Он заключается в том, что аллельные гены в гетерозиготном состоянии не сливаются, не разбавляются и не меняют друг друга. В результате мейоза в каждой гамете оказывается лишь одна из гомологичных хромосом, а следовательно, только один аллель гена («чистым» от другого аллеля).
Новаторской особенностью работы Менделя является статистическое мышление. Когда речь идет о гибридах второго поколения (II и III законы Менделя), невозможно однозначно предсказать, какие признаки будет иметь та или иная особь. Законы Менделя задают соотношение частот, которые характерны для особей с теми или иными признаками. Это связано с тем, что при формировании гамет у гетерозигот и при комбинировании гамет при оплодотворении ключевую роль играют случайные, вероятностные процессы.
Некоторые вопросы менделевской генетики
Анализирующее скрещивание. В случае полного доминирования для особи, в фенотипе которой проявился доминирующий признак, нельзя определить характер ее генотипа без дополнительных исследований. Наиболее эффективным способом анализа генотипа такой особи является анализирующее скрещивание, то есть скрещивание с особью, гомозиготной по рецессивному аллелю.
При моногибридном скрещивании может быть реализован один из следующих вариантов.

Если в потомстве появятся особи с рецессивным признаком в фенотипе, это означает, что исследуемая особь была гетерозиготной.
В ряде случаев наблюдается более сложный характер наследования признаков, чем зарегистрирован в экспериментах Менделя. Он может быть связан с взаимодействием генов. Так, в случае взаимодействия двух аллелей одной пары, кроме доминирования может наблюдаться неполное доминирование (промежуточный характер состояния признаков у гетерозигот). кодоминирование (проявление одновременно доминантного и рецессивного состояний признаков в гетерозиготе) и сверхдоминирование (более сильное проявление доминантного признака у гетерозигот, чем у гомозигот). Примерами взаимодействия генов из разных пар могут быть комплементарное действие (взаимное дополнение), эпистаз (подавление) и полимерия (множественное действие).
Неполное доминирование. В ряде случаев для других видов и признаков гетерозиготы имеют собственный фенотип, промежуточный между фенотипами гомозигот.
Так, при скрещивании двух форм душистого горошка, отличающихся по окраске цветов (А — красный, а — белый), гибриды (Аа) имеют промежуточную розовую окраску. Во втором поколении расщепление по фенотипу отвечает расщеплению по генотипу 1 : 2 : 1.
Кодоминирование характеризуется таким же расщеплением, как и неполное доминирование, но отличается от него тем, что у гетерозигот проявляются состояния признаков, связанные с действием аллелей. Например, каждый из аллельных генов кодирует определенный белок, и у гетерозиготного организма синтезируются они оба. В таких случаях путем биохимического исследования можно установить гетерозиготность без проведения анализирующего скрещивания. Примером такого наследования являются группы крови по системе АВ0.
У человека зарегистрированы четыре разные группы крови системы АВ0, отличающиеся по составу антигенов на поверхности эритроцитов и антител сыворотки крови. Группа крови I (0) характеризуется отсутствием в эритроцитах антигенов А и В и наличием в сыворотке крови антител α и β, группа крови II (А) связана с содержанием антигена и антител β, группа крови III (В) — антигена и антител α, группа крови IV (АВ) — антигенов А и В и отсутствием антител.
Установлено, что наследование групп крови человека связано с тремя аллелями одного гена (IA, Iв, i). При этом I (0) группа обусловлена рецессивным аллелем i, над которым доминируют как аллель IA, который определяет П группу, так и аллель Iв, от которого зависит III группа. Эти аллели кодоминантные, и встречаясь в гетерозиготе, определяют IV группу. Таким образом, I группа крови бывает лишь при генотипе ii, II — при генотипах IAIA и IAi, III — при генотипах IвIв и IBi, IV — при генотипе IАIВ.
Летальные аллели — аллельные гены, которые, проявляясь в фенотипе, влекут за собой гибель организма на той или иной стадии его развития. Чаще всего являются рецессивными и поэтому могут проявляться в фенотипе только в гомозиготном состоянии. Подобный тип наследования известен для серых каракулевых овец, платинового цвета лис и др.
В ряде случаев расщепление во втором поколении отличается от ожидаемого в связи с тем, что зиготы, содержащие определенные генотипы, оказываются нежизнеспособными. Примером служит скрещивание черных и желтых мышей, при котором в потомстве появляются индивиды этих цветов в соотношении 1 : 1. При скрещивании черных особей между собой в потомстве появляются лишь черные особи. А при скрещивании желтых — расщепление 2 : 1. Такое наследование желтого цвета обусловлено доминантным аллелем гена, но эта окраска реализуется лишь в гетерозиготном состоянии. Гомозиготные особи по доминантному аллелю погибают еще в эмбриональном состоянии.
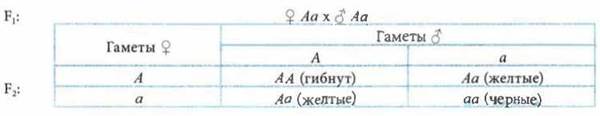
Генотип как целостная исторически сложившаяся система
Многие из рассмотренных выше признаков обусловлены одной парой генов. Кроме этих случаев, существует огромное количество свойств и признаков животных и растений, которые определяются двумя, тремя или даже многими парами генов. Изучение наследования таких признаков дало основание сформулировать положение о взаимодействии генов: не одна пара, а взаимодействие нескольких пар генов определяют, какой признак разовьется в организме.
Так, путем гибридологического анализа установлено, что форма гребня кур определяется двумя парами генов, молочная продуктивность крупного рогатого скота или морозостойкость злаков — многими парами генов. Вместе с тем оказалось, что одна и та же пара генов может содержать генетическую информацию о нескольких различных признаках организма (множественное действие генов). Например, у большинства растений с красными цветками в стеблях тоже есть пигмент. У дрозофилы ген, определяющий отсутствие пигментов в глазах, снижает плодовитость, влияет на окраску некоторых внутренних органов и уменьшает продолжительность жизни, то есть ген проявляет множественное действие.
Эти наблюдения углубляют представления о генотипе. Факт расщепления в потомстве гибридов позволяет утверждать, что генотип состоит из отдельных элементов — генов, которые могут наследоваться независимо. В то же время генотип не может рассматриваться как простая механическая сумма отдельных генов.
Генотип любого организма представляет собой сложную целостную систему взаимодействующих генов. Эта целостность генотипа возникла исторически в процессе эволюции вида. Она выражается, прежде всего, в тесном взаимодействии друг с другом отдельных генов. Какой фенотип разовьется на основе такого генотипа, будет зависеть как от самого генотипа, так и от тех условий, в которых развивается данный организм.
Изучение генотипов пород сельскохозяйственных животных и сортов растений — необходимая предпосылка для селекционной работы. Зная генетическую обусловленность тех или иных признаков, можно заранее спланировать такие схемы скрещиваний, которые обеспечат получение нужного потомства.
Хромосомная теория наследственности
Результаты исследований Т. Х. Моргана и его сотрудников (1911-1920 гг.) легли в основу предложенной хромосомной теории наследственности, которая во многом определила развитие генетики и биологии в целом. Она позволила выяснить материальную основу законов наследственности, исследованных Г. Менделем, и установить, почему при определенных случаях наследование признаков от них отклоняется.
Основные положения хромосомной теории наследственности
1. Гены находятся в хромосомах. Каждая хромосома представляет собой группу сцепления генов. Количество групп сцепления каждого вида равна гаплоидному набору хромосом (у дрозофилы — 4, гороха — 7, человека — 23 и т. д.).
2. Каждый ген в хромосоме занимает определенное место (локус).
3. Гены в хромосомах расположены линейно.
4. Расстояние между генами в хромосоме пропорционально проценту кроссинговера между ними.
5. Независимое расщепление генов происходит, если они находятся в разных гомологичных хромосомах.
6. Отдельное наследование генов одной группы сцепления может происходить при кроссинговере.
Гены, находящиеся в одной хромосоме, сцеплены не абсолютно. Во время профазы мейоза, при конъюгации хромосом гомологичные хромосомы способны обмениваться идентичными участками. Этот процесс получил название кроссинговер. Он может происходить в любом участке хромосомы, даже в нескольких местах одной хромосомы. Чем дальше друг от друга расположены локусы в одной хромосоме, тем чаще между ними следует ожидать перекрест и обмен участками. Этот процесс имеет большое значение для эволюции, потому что значительно увеличивает возможности изменчивости. Отбор в процессе эволюции происходит не целыми группами сцепления, а группами генов и даже отдельными генами.
Примером тесного сцепления генов у человека может служить наследование резус-фактора (определяется тремя рядом расположенными генами), гемофилии и дальтонизма в Х-хромосоме.
Генетические карты хромосом
Существование кроссинговера позволило школе Моргана в 1911-1914 гг. разработать метод построения генетических карт хромосом. В основу метода положено представление о расположении генов по длине хромосомы в линейном порядке. За единицу расстояния между двумя генами принято принимать 1% перекреста между ними. Генетические карты строятся на основе гибридологического анализа.
Предположим, что к одной группе сцепления принадлежат гены А и В. Между ними обнаружен кроссинговер в 10%. Итак, гены А и В находятся на расстоянии 10 единиц. К этой же группе сцепления принадлежит ген С. Чтобы узнать его место в хромосоме, необходимо выяснить, какой процент перекреста он дает с каждым из двух генов.
Например, если с геном А он дает 3%, то можно предположить, что ген С находится или между А и В, или с противоположной стороны, то есть А расположен между С и В. Если процент кроссинговера между С и В окажется 7%, то первое предположение подтверждается, если же перекрест составит 13%, то второе. Наиболее подробные карты хромосом составлены для дрозофилы — классического генетического объекта. Из растений сравнительно хорошо в этом отношении изучены кукуруза и томаты, из животных — куры и мыши. Практически полностью составлены генетические карты и для человека.
Генетика пола
Хромосомная теория наследственности позволила ответить на вопрос, чем определяется пол у разнополых животных и как достигается равенство числа особей обоих полов в каждом поколении. Хромосомный набор самцов и самок большинства разнополых организмов неодинаков: в нем существуют два типа хромосом. Это аутосомы — хромосомы, одинаковые у обоих полов, и половые хромосомы (гетерохромосомы), те, по которым самцы и самки отличаются друг от друга.
В клетках тела организма обычно находятся две половые хромосомы. Если пара половых хромосом представлена двумя одинаковыми хромосомами (их называют Х-хромосомами), особь называется гомогаметной, если двумя разными — Х- и Y-хромосомами — гетерогаметной.
У большинства животных, в том числе у человека, женскому полу соответствует набор половых хромосом ХХ, мужскому — ХY. У рептилий, птиц и ряда видов бабочек женские особи имеют разные половые хромосомы, а мужские — одинаковые. У некоторых насекомых (прямокрылые) Y-хромосома отсутствует. В таком случае кариотип самцов 2n + ХО, самок — 2n + ХХ.
Наследование, сцепленное с полом
Признаки, которые наследуются через половые хромосомы, получили название сцепленных с полом. У человека признаки, унаследованные через Y-хромосому, могут быть только у мужчин; а унаследованные через Х-хромосому — у представителей обоих полов. Женщина может быть как гомо-, так и гетерозиготной по генам, локализованным в Х-хромосоме, а рецессивные аллели генов выявляются только в гомозиготном состоянии. Поскольку у лиц мужского пола только одна Х-хромо- сома, все локализованые в ней гены, даже рецессивные, сразу же проявляются в фенотипе. Такой организм называют гемизиготным. При записи схемы передачи признаков, сцепленных с полом, в генетических формулах, наряду с символами генов, указывают и половые хромосомы.
У человека некоторые патологические состояния наследуются сцеплено с полом (гемофилия). Если рецессивные признаки, унаследованные через Х-хромосому у женщин, проявляются только в гомозиготном состоянии, то доминантные в одинаковой степени проявляются у обоих полов. К таким признакам у человека относятся витаминустойчивый рахит, темная эмаль зубов и др. Признаки, наследуемые через Y-хромосому, называются голандрическими. Пример: у человека интенсивное развитие волос на краю ушной раковины.
Организация генома у различных групп организмов
Генетический материал всех живых организмов организован в специальные структуры — хромосомы, представленные в многоклеточных организмах особыми тельцами, которые специфически окрашены специальными красителями и поэтому их хорошо видно при микроскопическом изучении клеток.
Генетический материал вирусов может включать от одного до нескольких сотен генов, практически все являются структурными.
Геном прокариотов представляет собой голую молекулу ДНК из двух цепочек — нуклеоид. Иногда эта молекула линейная, но в основном она представлена кольцевыми формами, которые образуются за счет соединения липких концов молекул. Геном прокариотов содержит как структурные, так и регуляторные гены.
Хромосомы эукариот содержат огромное количество структурных и функциональных генов. Геном дрозофилы содержит 10 000 структурных генов, а человека — 30 000. Вещество хромосом эукариот — хроматин — представляет собой сложно организованные нуклеопротеидные комплексы, включающие ДНК и два вида белков: гистоны и кислые белки. Молекула ДНК в каждой хромосоме одна, она непрерывна. Ее длина в развернутом виде может достигать десятков и сотен сантиметров.
Размещение молекулы ДНК в ядре эукариотической клетки достигается ее компактизацией. Это происходит благодаря взаимодействию ДНК и белков. На первом этапе спирализации ДНК, как сильная кислота, связывается с основными белками — гистонами. Кроме того, эти белки играют важную роль в регуляции активности генов. При этом двойная спираль ДНК наматывается на молекулы гистонов, они, в свою очередь, составляют своеобразную сердцевину, и размер ДНК уменьшается почти в пять раз. Образуются нуклеосомы.
При вторичной спирализации образуется хроматида толщиной около 25 нм. Нуклеосомная нить взаимодействует с другими белками, образуется спираль более высокого уровня. При заключении в хромосому каждая из двух хроматид еще раз дополнительно спирализуется. Такая упорядоченность обеспечивает их сохранность, четкое и правильное распределение хромосом при делении клеток. Препятствие транскрипции преодолевается деспирализацией в интерфазе, когда хромосомы приобретают толщину 25 нм.
Цитоплазматическая наследственность
Отдельные органеллы цитоплазмы (митохондрии, пластиды) имеют собственную ДНК, а также собственную систему синтеза белка. Генетическая информация в этом случае передается вместе с цитоплазмой, при этом она получила название цитоплазматической (внеядерной, внехромосомной).
Установлено, что собственную ДНК имеют пластиды (пластидная ДНК), митохондрии (митохондриальная ДНК) и некоторые другие органеллы. Эти цитоплазматические структуры способны к саморазмножению. Именно с ними связана передача цитоплазматической наследственности. Тем не менее, проявление этой формы наследственности находится под контролем ядерной ДНК.
Явление цитоплазматической наследственности было открыто одним из основателей генетики К. Коренсом.